

|
Date
|
Mercredi 03 avril 2024 |
Matière
|
SVT_THOUAND 228
|
|
Chapitre
|
6.1 / 6.2
|
Séquence
|
|
|
Résumé
|
Cours 6.1 La cellule musculaire [suite]
IV Les myopathies
Les myopathies sont des maladies génétiques qui se caractérisent par une dégénérescence des cellules musculaires.Analyse des documents pages 430-431 Le gène DMD code pour la dystrophine, une protéine qui relie les fibres du cytosquelette (l’ actine notamment) aux fibres de collagène de la matrice extracellulaire, via des protéines membranaires. matrice extracellulaire : réseau de longues molécules, notamment de collagène chez les animaux, formant un gel entre les cellules qui assure leur cohésion. La myopathie de Duchenne (DMD) est une maladie génétique due à une mutation, récessive, affectant le gène DMD porté par le chromosome X (d’où le fait que seuls les garçons soient touchés). Selon la mutation, la dystrophine est absente ou anormale. Il en résulte une absence de lien entre le cytosquelette des cellules musculaires et la matrice extracellulaire. Au fur et à mesure des cycles de contraction/relâchement des myofibrilles, cette absence de lien avec la matrice extracellulaire engendre une rupture de la membrane plasmique, qui provoque la mort de la fibre musculaire. Si au début de la vie les fibres musculaires sont régénérées, cela finit par ne plus être le cas : elles deviennent de moins en moins nombreuses, au profit de la matrice extracellulaire qui envahit le muscle. Les muscles finissent ainsi par ne plus être fonctionnels. En conclusion, les myopathies sont dues à un défaut dans les interactions entre cytosquelette, protéines membranaires et matrice extracellulaire. Chapitre 6.2 L’origine de l’ATP
Tous les processus cellulaires nécessitant de l’énergie reposent sur l’hydrolyse de l’ATP (cf. chapitre 6.1) : l’ATP doit donc être constamment produit afin que la cellule reste en vie.Rappels Oxyder = prendre des électrons La matière organique (ex. [latex]\mathrm{CH_4}[/latex])est formée de carbone réduit. Le [latex]\mathrm{CO_2}[/latex] représente au contraire l’état le plus oxydé du carbone. Sur Terre, les conditions de vie sont oxydantes : par conséquent, l’oxydation de la matière organique libère de l’énergie. La production d’ATP repose sur un couplage énergétique avec des réactions d’ oxydation de matière organique (le glucose). RESPIRATION CELLULAIRE : Si l’oxydation du glucose est totale ( [latex]\mathrm{C_6H_{12}O_6 \rightarrow CO_2}[/latex]), l’énergie libérée est importante, ce qui permet de produire beaucoup d’ATP ( 36 moles par mole de glucose). FERMENTATIONS : Si l’oxydation du glucose est partielle (= le produit de la réaction est encore une molécule organique, et non du [latex]\mathrm{CO_2}[/latex]), la faible quantité d’énergie libérée ne permet de synthétiser qu’une petite quantité d’ATP. I La respiration cellulaire
I.1 Vue d’ensemble
paragraphe de présentation avant de rentrer dans le détail : toutes les idées de cette partie I.1 doivent être claires afin de comprendre et apprendre les différentes étapes de la respiration cellulaire I.1.a Bilan
La respiration cellulaire est une oxydation totale du glucose ( [latex]\mathrm{\mathrm{C_6H_{12}O_6\to6CO_2}}[/latex]), qui permet de synthétiser 36 moles d’ATP pour 1 mole de glucose. I.1.b Étapes
La respiration cellulaire se déroule en 3 étapes.Les deux premières étapes conduisent à l’ oxydation du glucose. Les électrons sont transférés à des molécules de [latex]\mathrm{\mathrm{NAD^+}}[/latex], qui sont ainsi réduites en [latex]\mathrm{\mathbf{NADH,H^+}}[/latex] (appelées composés réduits, ou pouvoir réducteur, et parfois notées [latex]\mathrm{\mathrm{\mathfrak{R}H_2}}[/latex]). La première étape est la glycolyse, elle a lieu dans le cytoplasme. Le glucose est oxydé en pyruvate (= acide pyruvique). La seconde étape est le cycle de Krebs, elle a lieu dans la matrice de la mitochondrie. Le pyruvate est oxydé en [latex]\mathrm{\mathrm{CO_2}}[/latex]. Lors de la troisième étape, les composés réduits [latex]\mathrm{\mathrm{NADH,H^+}}[/latex] sont réoxydés en [latex]\mathrm{\mathbf{NAD^+}}[/latex] ; leurs électrons sont cédés au [latex]\mathrm{\mathrm{O_2}}[/latex]). Ce transfert d’électrons permet la synthèse d’une grande quantité d’ATP. Cette troisième étape se déroule au niveau de la chaîne respiratoire mitochondriale, dans la membrane interne des mitochondries. I.1.c Rappels sur la structure de la mitochondrie
Schéma d’une mitochondriemembrane externe, espace intermembranaire, membrane interne avec crêtes mitochondriales, matrice I.2 Déroulement de la respiration cellulaire
I.2.a Première étape : la glycolyse
C’est une suite de réactions qui ont lieu dans le cytoplasme :[latex]\mathrm{\mathbf{glucose\to2\,pyruvate}}[/latex] Les réactions de la glycolyse sont couplées à la synthèse de 2 ATP et de 2 [latex]\mathrm{\mathbf{NADH,H^+}}[/latex]. I.2.b Deuxième étape : le cycle de Krebs
C’est une suite de réactions qui ont lieu dans la matrice de la mitochondrie.Une des premières réactions fait intervenir une molécule, qui est reformée lors de la dernière réaction : on parle donc de cycle. [latex]\mathrm{\mathbf{2\,pyruvate\to6\,CO_2}}[/latex] Les réactions du cycle de Krebs sont couplées à la synthèse de 2 ATP et de 10 [latex]\mathrm{\mathbf{NADH,H^+}}[/latex]. I.2.c Troisième étape : production d’ATP par réoxydation des composés réduits au niveau de la chaîne respiratoire |
||
|
À voir
|
|||
|
À faire
pour le
05-04-2024
|
DM sur l’ [latex]\mathrm{\alpha}[/latex]bungarotoxine
Test de connaissances sur le thème 6 |
||
|
Commentaires
|
|||
|
Date
|
Vendredi 22 mars 2024 |
Matière
|
SVT_THOUAND 225
|
|
Chapitre
|
5.1/5.2
|
Séquence
|
|
|
Résumé
|
Cours IV La synapse neuro-musculaire [suite]
À retenir :— neurotransmetteur = acétylcholine — la fixation d’acétylcholine sur ses récepteurs membranaires génère une dépolarisation de la membrane plasmique de la cellule musculaire, qui se propage le long de la cellule jusqu’aux tubules T : potentiel d’action musculaire — => ce potentiel d’action musculaire entraîne l’ augmentation de la concentration cytoplasmique en [latex]\mathrm{Ca^{2+}}[/latex] (par ouverture de canaux calciques voltage-dépendants dans la membrane du réticulum sarcoplasmique) — => contraction de la fibre musculaire Chapitre 5.2 Cerveau et mouvement volontaire
I Les cellules gliales
Les cellules du cerveau sont :— les neurones, qui transmettent les messages nerveux — les cellules gliales (exemples : astrocytes, oligodendrocytes), qui assurent différentes fonctions (nutrition, immunité, etc.) permettant le bon fonctionnement du cerveau II Les aires motrices
C’est dans l’ aire motrice primaire que se trouvent les corps cellulaires des neurones pyramidaux qui envoient des messages nerveux aux motoneurones afin de réaliser des mouvements volontaires.Les neurones pyramidaux élaborent un message nerveux à partir des informations reçues par les aires motrices secondaires. Les fibres nerveuses des neurones pyramidaux forment des faisceaux dans la moelle épinière, jusqu’aux corps cellulaire des motoneurones (synapse cholinergique). Une lésion de la moelle épinière entraîne ainsi une paralysie de la partie du corps inférieure à la lésion (para- ou tétra-plégie). III Intégration des différents messages par le neurone moteur
intégration d’informations = production d’un message unique en fonction des différentes informations reçuesAu niveau du corps cellulaire d’un neurone : — les neurotransmetteurs excitateurs provoquent une dépolarisation — les neurotransmetteurs inhibiteurs provoquent une hyperpolarisation Si la dépolarisation au niveau du corps cellulaire dépasse un certain seuil, des potentiels d’action sont générés au niveau de l’axone du neurone. Au niveau du corps cellulaire d’un neurone, les modifications du potentiel membranaire (dépolarisation ou hyperpolarisation) s’ajoutent au cours du temps = sommation temporelle. Ainsi, une succession de petites stimulations rapprochées peut générer des potentiels d’action dans l’axone. Par ailleurs, les effets des différentes synapses reliés au corps cellulaire s’ajoutent également = sommation spatiale. Ainsi, les stimulations conjointes par plusieurs synapses excitatrices peuvent générer des potentiels d’action dans le neurone ; inversement, l’action inhibitrice de certaines synapses peut empêcher la dépolarisation due aux synapses excitatrices de dépasser le seuil de génération de PA. |
||
|
À voir
|
Plus de détails sur la contraction musculaire et les synapses neuro-musculaires |
||
|
À faire
pour le
28-03-2024
|
Réaliser les deux sujets d’exercice 2 distribués en classe (botox/benzodiazépines).
Pour gagner du temps, vous pouvez les rédiger de manière très condensée. Ce qui compte est de montrer que :
— vous avez extrait l’information qu’apporte chaque document
— vous avez les connaissances nécessaires pour utiliser ces informations
— que vous êtes capable d’enchaîner logiquement toutes ces informations dans un argumentaire cohérent répondant au problème
|
||
|
Commentaires
|
|||
|
Date
|
Jeudi 21 mars 2024 |
Matière
|
SVT_THOUAND 223
|
|
Chapitre
|
5.1 Les réflexes
|
Séquence
|
|
|
Résumé
|
Cours III.2 Transmission des messages nerveux au niveau d’une synapse
Schéma de la transmission des messages nerveux au niveau d’une synapse neuro-neuroniqueAU niveau d’une synapse, le message nerveux est de nature chimique et est codé en concentration de neurotransmetteurs. Un neurone ne produit qu’un seul type de neurotransmetteur (= neuromédiateur) ; par conséquent, une synapse ne fonctionne qu’avec un seul neurotransmetteur. Exemples : — synapse à acétylcholine (= synapse cholinergique) — synapse à dopamine (= dopaminergique) Dans l’arc-réfexe du réflexe myotatique, le neurotransmetteur est l’ acétylcholine. Synapses inhibitrices : Dans ce cas, la fixation du neurotransmetteur sur son récepteur inhibe (empêche) la formation de potentiels d’action dans le neurone post-synaptique. Exemple : — synapse à GABA (le GABA est un neurotransmetteur inhibiteur) Remarque Différentes substances peuvent altérer le fonctionnement des synapses : — en se fixant sur les récepteurs, et en mimant la fixation du neurotransmetteur — en se fixant sur les récepteurs, sans avoir d’action, mais en empêchant la fixation du neurotransmetteur — en augmentant l’effet de la fixation du neurotransmetteur sur son récepteur — en empêchant la dégradation du neurotransmetteur IV La synapse neuro-musculaire
= synapse entre neurone moteur et cellule musculaireSchéma fonctionnel de la synapse neuro-musculaire Évaluation Test de connaissances |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 20 mars 2024 |
Matière
|
SVT_THOUAND 222
|
|
Chapitre
|
5.1 Les réflexes
|
Séquence
|
|
|
Résumé
|
Cours Retour sur le chapitre 3.4
Analyse des documents 5 à 7 page 273Mise en évidence des conséquences d’une faible diversité génétique dans une population (dans le cas de la bananier, due à la culture d’une seule variété, formée de clones) sur la sensibilité aux maladies, et de la nécessité de conserver de la diversité génétique afin d’avoir des réserves génétiques de caractères de résistances. Chapitre 5.1
II Messages nerveux et potentiel d’action [suite]
II.2 La propagation des potentiels d’action
Un potentiel d’action est une variation transitoire du potentiel de membrane, qui n’est que locale.Un potentiel d’action est capable de se propager le long d’une fibre nerveuse, à une vitesse d’1 m/s. Les fibres nerveuses sont très souvent myélinisées (c’est-à-dire recouvertes d’une gaine de myéline formée par des cellules de Schwann séparées par des nœuds de Ranvier) : cela permet que la vitesse de propagation des potentiels d’action soit de 100 m/s. NB : la sclérose en plaques est une maladie auto-immune qui s’attaque à cette gaine de myéline. II.3 Le message nerveux : codage en fréquence de potentiels d’action
Au niveau d’une fibre nerveuse, un message nerveux est formé par un train de potentiels d’action.Un message nerveux est un message de nature électrique (PA = variations du potentiel membranaire), dont est codée par la fréquence des potentiels d’action. III Transmission des messages nerveux au niveau des synapses
Les potentiels d’action étant des variations du potentiel de membrane, ils ne peuvent pas être transmis d’un neurone à l’autre au niveau d’une synapse (car il n’y a pas de continuité des membranes). III.1 Structure des synapses
À connaître :— neurone présynaptique, fente synaptique, neurone postsynaptique — bouton synaptique (extrémité du neurone présynaptique), vésicules remplies de molécules de neurotransmetteur ; récepteurs au neurotransmetteur (molécules portées par la membrane du neurone postsynaptique) Schéma d’une synapse neuro-neuronique |
||
|
À voir
|
|||
|
À faire
pour le
21-03-2024
|
Jeudi 21 mars
Court test de connaissances sur : — chapitre 5.1 — chapitre 5.2 (cf. poly distribué la semaine dernière, et disponible ci-dessus en couleurs) — questions sur l’émission de radio (cf. travail donné vendredi dernier : je vous poserai quelques unes des questions posées parmi celles que vous mises sur le bloc-note collaboratif) |
||
|
Commentaires
|
|||
|
Date
|
Vendredi 15 mars 2024 |
Matière
|
SVT_THOUAND 214
|
|
À voir
|
Émission de radio sur les NTG : lien alternatif sur le peertube de louisematic Courte (8 mn) émission de radio présentant ce que sont les nouvelles techniques génomiques (NTG) |
||
|
À faire
pour le
21-03-2024
|
Écouter l’émission de radio suivante : https://www.radiofrance.fr/franceculture/podcasts/la-science-cqfd/ogm-ngt-de-la-bonne-ou-de-la-mauvaise-graine-6391523
Posez une question portant sur cette émission, dans le bloc-note collaboratif https://louisematic.site/pad/p/ngt-3x38qdm |
||
|
Commentaires
|
|||
|
Date
|
Vendredi 15 mars 2024 |
Matière
|
SVT_THOUAND 215
|
|
À voir
|
Émission de radio sur les NTG : lien alternatif sur le peertube de louisematic Courte (8 mn) émission de radio présentant ce que sont les nouvelles techniques génomiques (NTG) |
||
|
À faire
pour le
21-03-2024
|
Écouter l’émission de radio suivante : https://www.radiofrance.fr/franceculture/podcasts/la-question-du-jour/quels-sont-les-risques-des-nouveaux-ogm-5788895
Posez une question portant sur cette émission, dans le bloc-note collaboratif https://louisematic.site/pad/p/ngt-3x38qdm |
||
|
Commentaires
|
|||
|
Date
|
Mercredi 24 janvier 2024 |
Matière
|
SVT_THOUAND 184
|
|
Chapitre
|
3.2 La plante, productrice de matière organique
|
Séquence
|
|
|
Résumé
|
Cours Chapitre 2 — La plante, productrice de matière organique
Introduction [suite]
Les plantes sont autotrophes grâce à la photosynthèse= succession de réactions biochimiques dont le bilan est : [latex]\mathrm{\mathbf{6CO_2 + 6H_2O \overset{+\ énergie\ lumineuse}{\rightarrow} \underset{glucose\ (énergie\ chimique)}{C_6H_{12}O_6} + 6O_2}}[/latex] Photosynthèse du point de vue du carbone : transformation du [latex]\mathrm{\mathrm{CO_2}}[/latex] en matière organique <=> réduction du [latex]\mathrm{\mathrm{CO_2}}[/latex] (à l’inverse, la respiration cellulaire est une oxydation totale de la matière organique, le [latex]\mathrm{\mathrm{CO_2}}[/latex] représentant l’état le plus oxydé d’un atome de C) La photosynthèse permet donc la fixation du carbone atmosphérique, c’est-à-dire son passage de l’atmosphère vers la biosphère sous forme de matière organique (cf. cours sur le climat). Photosynthèse du point de vue énergétique : l’ énergie lumineuse est convertie en énergie chimique stockée dans la matière organique (glucose et autres molécules organiques qui en dérivent – cf. II.2) I La photosynthèse
Se déroule dans les chloroplastesSchéma d’un chloroplaste [pas exigible] : 2 membranes entourant le stroma ; compartiments aplatis = thylakoïdes, dont le membrane contient de nombreux pigments (= molécules colorées = molécules absorbant de la lumière) I.1 Première phase : phase claire = phase photochimique
Une molécule de chlorophylle a absorbe la lumière, ce qui lui fait perdre un électron, qui est transmis à d’autres molécules. L’énergie lumineuse est ainsi convertie en énergie chimique, contenue dans les molécules produites : ATP et pouvoir réducteur. Ces molécules, riches en énergie et en électrons (= réduites), serviront dans la seconde phase pour réduire le [latex]\mathrm{\mathrm{CO_2}}[/latex] en glucose. La chlorophylle récupère cet électron par photolyse de l’eau : [latex]\mathrm{\mathrm{2H_2O \rightarrow O_2 + 4H^+ + 4e^-}}[/latex] N.B. : Le « pouvoir réducteur » désigne des molécules de [latex]\mathrm{NADPH + H^+}[/latex] (nicotinamide adénine dinucléotide phosphate), parfois aussi notées [latex]\mathrm{\Re H_2}[/latex] Schéma-bilan de la phase claire (avec le nom des molécules chargées en énergie et en électrons qui sont produites : ATP et NADPH,H+ — noms qui ne sont pas exigibles mais que vous pouvez rencontrer) Les autres pigments chlorophylliens (chlorophylle b, caroténoïdes, etc.) sont également capables d’absorber la lumière ; ils transmettent ensuite cette énergie à la chlorophylle a, seul pigment capable de perdre un électron. Ces autres pigments n’absorbent pas les mêmes longueurs d’onde que la chlorophylle a (d’où leur couleur différente) ⇔ ils n’ont pas le même spectre d’absorption (cf. p. 223 doc. 4 : par ex. les caroténoïdes absorbent plutôt vers 500 nm — d’où leur couleur rouge orangé). Ils permettent donc à la photosynthèse d’être réalisée même à partir de longueurs d’onde qui ne sont pas absorbées par la chlorophylle a ⇔ ils élargissent le spectre d’action de la photosynthèse (longueurs d’onde permettant d’effectuer la photosynthèse : cf. courbe « activité photosynthétique » p. 223 doc. 4). II.2 Seconde phase : phase sombre = phase chimique
Les molécules produites lors de la phase claire, [latex]\mathrm{\mathbf{ATP et NADPH+H^+}}[/latex] (riches en énergie et en électrons) permettent la réduction du CO2 en glucose par une succession cyclique de réactions : le cycle de Calvin - Benson.La réaction permettant de fixer le [latex]\mathrm{CO_2}[/latex] dans la matière organique est catalysée par une enzyme : la RubisCo. On parle de cycle car les molécules consommées lors de la première réaction sont produites par les dernières réactions (elles n’apparaissent donc pas dans le bilan). Cette phase ne nécessite pas de lumière (l’énergie lumineuse a été convertie lors de la première phase en énergie chimique contenue dans l’ [latex]\mathrm{ATP et NADPH+H^+}[/latex]). Schéma-bilan de la phase sombre : cycle de Calvin |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 12 janvier 2024 |
Matière
|
SVT_THOUAND 166
|
|
Chapitre
|
3.1 L’organisation fonctionnelle des plantes à fleurs
|
Séquence
|
|
|
Résumé
|
Cours / TD I.1 Feuilles [suite]
Exemple pour un arbre (de 2 t) : surface foliaire = [latex]\mathrm{\mathbf{400 m^2}}[/latex] Exercice — Calcul de la surface d’échange foliaire d’un végétal par rapport à sa masse (S/m) Données : la surface foliaire d’un arbre de 2 t est S = 400 [latex]\mathrm{m^2}[/latex] Conversions : 2 t = 2 000 kg ; 400 [latex]\mathrm{m^2}[/latex] = 4 000 000 [latex]\mathrm{cm^2}[/latex] Résultats : S/m = 0,2 [latex]\mathrm{\mathbf{m^2.kg^{-1}}}[/latex] = 2000 [latex]\mathrm{cm^2.kg^{-1}}[/latex] I.2 Racines
Fonction principale : absorption de l’eau et des sels minéraux du sol (+ ancrage dans le sol)Très grande surface d’échange entre le sol et le végétal car [cf. p. 201] : — racines = structures longues, fines, très ramifiées (nombreuses) — présence à l’extrémité des racines de nombreux poils absorbants = cellules racinaires allongées, pénétrant dans le sol où elles prélèvent l’eau et les sels minéraux (c’est uniquement à ce niveau que se réalise l’absorption, pas au niveau de l’épiderme) Schéma de l’extrémité d’une racine en vue longitudinale Schéma d’une coupe transversale théorique de racine Exercices : — Calcul du rapport S/m pour les racines d’une Arabidopsis thaliana : Données : S = 0,81 cm² ; m = 1,6 mg Résultat : S/m = 51 [latex]\mathrm{\mathbf{m^2.kg^{-1}}}[/latex] N.B. : ceci ne concerne que la surface des racines, mais pas la surface d’absorption (c’est la surface de l’épiderme racinaire qui est prise en compte, pas celle des poils absorbants) — Calcul de la surface totale des poils absorbants (surface d’échange) d’une céréale (seigle) : Données : - poil absorbant : diamètre = 14 [latex]\mathrm{\mu m}[/latex] ; longueur = 0,7 mm - nombre de poils absorbants : 14 milliards => surface d’un poil absorbant : [latex]\mathrm{\mathrm{S_{poil} = 7.10^{-4}\times\Pi\times14.10^{-6}}}[/latex] (en [latex]\mathrm{m^2}[/latex]) => surface totale d’échange : [latex]\mathrm{\mathrm{S_{tot} = 7.10^{-4}\times\Pi\times14.10^{-6}\times14.10^9}}[/latex] => S = 415 m² (la surface d’échange entre les racines du seigle et le sol est supérieure à la surface d’un terrain de tennis !) Mycorhizes
L’absorption de l’eau et des sels minéraux est considérablement favorisée par des symbioses avec des champignons = mycorhizes, dont les filaments mycéliens sont en contact étroit avec les racines et pénètrent dans le sol, augmentant encore la surface d’échange avec le sol.C’est une symbiose car : – association étroite (les mycorhizes pénètrent dans les racines, voire parfois dans les cellules racinaires) – à bénéfice mutuel : en retour, le végétal fournit au champignon de la matière organique, produite par photosynthèse II Les tissus conducteurs
Deux réseaux parallèles permettent les échanges de matière entre organes aériens et souterrains II.1 La sève brute : le xylème
– = eau et sels minéraux– ascendante (racines -> feuilles) – transportée par les vaisseaux du xylème = cellules allongées, mortes, dont les parois épaisses forment des tubes connectés (leur paroi cellulosique est enrichie en lignine). L’ensemble de ces vaisseaux forment un tissu nommé xylème (= bois). |
||
|
À voir
|
Article « Les racines, grandes oubliées de l’agronomie et clés d’un futur écoresponsable » Article « Un atlas des systèmes racinaires sauvé de l’oubli et accessible en ligne » |
||
|
À faire
|
|
||
|
Commentaires
|
|||

|
Date
|
Jeudi 11 janvier 2024 |
Matière
|
SVT_THOUAND 167
|
|
Chapitre
|
3.1 L’organisation fonctionnelle des plantes à fleurs
|
Séquence
|
|
|
Résumé
|
Cours Thème 3 — De la plante sauvage à la plante domestiquée
Chapitre 1 — L’organisation fonctionnelle des plantes à fleurs
Plantes à fleurs (= angiospermes) : le plus grand nombre des végétaux [sont exclues de l’étude : les bryophytes (mousses), les ptéridophytes (fougères), les gymnospermes (conifères, ginkgo)] Organes végétatifs (non reproducteurs) d’une plante : racines, tiges, feuilles Mode de vie : vie fixée, à l’ interface sol/air, dans un environnement variable => contraintes : - contraintes trophiques (pas de possibilité de se déplacer vers des ressources de nutriments, d’où nécessité d’optimiser l’extraction des nutriments de son environnement proche) - contraintes reproductives (pb de la rencontre des gamètes entre individus qui ne peuvent s’approcher ; pb de la dissémination des descendants) - contraintes vis-à-vis des prédateurs (fuite impossible) - risque de dessiccation (milieu aérien très déshydratant) - environnement variable : variations journalières (pas de lumière la nuit, risque de déshydratation aux heures les plus chaudes), variations saisonnières (saison froide : risque de gel, peu de lumière / saison sèche) –I Des surfaces d’échanges de grande dimension
––I.1 Feuilles
Fonction principale : photosynthèse (=> besoin de lumière, eau, dioxyde de carbone) Schéma d’une coupe transversale de feuille – cellules photosynthétiques ( parenchyme palissadique) situées en face supérieure, du côté de la lumière – échanges gazeux permis par les stomates, et les lacunes du parenchyme lacuneux en face inférieure – stomate = orifice situé en face inférieure, dont l’ouverture peut être contrôlée (les deux cellules de garde peuvent fermer l’ostiole = ouverture entre ces deux cellules) => Échanges favorisés par : – grande surface, grâce à une structure aplatie et fine – stomates Problème : Favoriser les échanges gazeux au niveau des feuilles favorise l’apport de dioxyde de carbone, mais entraîne la perte d’eau par évapotranspiration. Différentes structures permettent de limiter ces pertes, notamment : — cuticule (couche imperméable) recouvrant la surface des feuilles (notamment la surface supérieure) — stomates situés en face inférieure (à l’ombre), pouvant se fermer si le risque de dessiccation est trop grand —présence possible de poils en face inférieure (emprisonnant une couche d’air) |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 10 janvier 2024 |
Matière
|
SVT_THOUAND 168
|
|
Chapitre
|
1.4 D’autres mécanismes contribuent à la diversité du vivant
|
Séquence
|
|
|
Résumé
|
Cours III Transmission culturelle
Chez certaines espèces animales, des comportements peuvent être acquis, par apprentissage avec imitation : on parle de transmission culturelle de caractères. Attention, tous les comportements ne sont pas transmis culturellement, la plupart sont génétiques. N.B. : opposition nature/culture = inné/acquis = génétique/environnemental La diversité des comportements acquis participe à la diversification phénotypique (phénotype étendu), sans modification génétique. Groupes d’êtres vivants principalement concernés : oiseaux (notamment les corvidés), mammifères (notamment les primates) Caractères souvent transmis culturellement : — chant (oiseaux) – outils — chez l’humain : de nombreux comportements (langue par exemple) Exemples — p. 104 doc. 1 Exemple de transmission culturelle d’un comportement par apprentissage avec imitation – brindille dans l’oreille dans une population de chimpanzé (cf. lien ci-dessous) — p. 104 doc. 3 lavage du blé dans une population de macaques du Japon — différentes espèces d’oiseaux de GB qui ont adopté le comportement consistant à percer l’opercule des bouteilles de lait déposées à la porte des maisons Ces comportements sont acquis, notamment grâce à l’ apprentissage des juvéniles (jeunes) par imitation des adultes => ils sont transmis entre contemporains de génération en génération : ils sont héréditaires. [remarque : héréditaire = transmis de parent à descendant ; un caractère peut être héréditaire sans être génétique — exemple de la langue parlée chez les humains, qui se transmet généralement des parents aux enfants] => ces caractères culturels peuvent subir une évolution similaire aux caractères génétiques (évolution culturelle) : — apparition fortuite (comme les mutations font apparaître par hasard un caractère génétique) — possibilité de (contre-)sélection : si un comportement procure un avantage reproductif, la proportion des individus ayant adopté ce comportement sera plus élevée à chaque génération (puisqu’il est transmis aux descendants, qui dans ce cas ont plus de descendants que les autres) — disparition par hasard possible (si l’effectif de la population est réduit) |
||
|
À voir
|
La diversité des comportements transmis culturellement, de la drosophile à l’humain
Casser des noix, une question de culture chez le chimpanzé
Deux vrais jumeaux élevés l'un chez les nazis, l'autre dans une famille juive |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 08 décembre 2023 |
Matière
|
SVT_THOUAND 148
|
|
Résumé
|
Amylase
http://didac.free.fr/bac_s_archive_2013-20/ts2009polynesie/3.htm Résistance aux insecticides http://didac.free.fr/bac_s_archive_2013-20/ts18polynesie/3.htm Espèce de dauphin http://didac.free.fr/bac_s_archive_2013-20/ts17amerique_s/3.htm Pouillot verdâtre http://didac.free.fr/bac_s_archive_2013-20/ts14amerique_n/3.htm Coévolution http://didac.free.fr/bac_s_archive_2013-20/ts19amerique_n/3.htm Pour grand oral : http://didac.free.fr/bac_s_archive_2013-20/ts16pondichery/3.htm |
||
|
À voir
|
Document partagé où s’inscrire pour l’étude des articles Mini-conférence de Pierre-Henri Gouyon Guide de compréhension de la mini-conférence de Pierre-Henri Gouyon |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 08 décembre 2023 |
Matière
|
SVT_THOUAND 149
|
|
Chapitre
|
1.3 L’inéluctable évolution des génomes au sein des populations
|
Séquence
|
|
|
À voir
|
Document partagé où s’inscrire pour l’étude des articles Mini-conférence de Pierre-Henri Gouyon Guide de compréhension de la mini-conférence de Pierre-Henri Gouyon |
||
|
À faire
pour le
13-12-2023
|
Pour mercredi 13 décembre :
— regarder la mini-conférence de Pierre-Henri Gouyon (lien ci-dessus) Cette mini-conférence d’un grand scientifique français est très riche. Les informations peuvent d’ailleurs être réutilisées dans différents chapitres de l’année, et être sources de sujets de grand oral. Pour vous aider à comprendre et retenir les notions importantes, vous pouvez vous inspirer du guide « Pistes de réflexion sur la mini-conférence » (il ne s’agit pas d’un DM à faire, mais je pourrai vous poser certaines questions en classe, pour évaluer ce que vous avez retenu). C’était un document réalisé pour des élèves de seconde, vous devriez y arriver (PS La référence à un certain Rayan ne concerne donc aucun d’entre vous). Étude d’articles : — choisir deux dossiers, en inscrivant son nom sur le document partagé (lien ci-dessus). Il faut étudier un dossier traitant de l’évolution, et sur espèce/spéciation Lisez les articles, et préparez-vous pour mercredi à répondre à des questions sur les articles de la partie espèce/spéciation |
||
|
Commentaires
|
|||
|
Date
|
Samedi 25 novembre 2023 |
Matière
|
SVT_THOUAND 138
|
|
Résumé
|
Bac blanc |
||
|
À voir
|
|||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 24 novembre 2023 |
Matière
|
SVT_THOUAND 137
|
|
Résumé
|
TD Sujet d’exercice 2 : Moustiques et diversification des génomes http://louise.michel.g.free.fr/upload/thouand_e-47c5d3a-Bac-2021-m-tropole-septembre9-21svtj1me3-exo2-moustiques-insecticides-.pdf Sujet d’exercice 2 : Histoire géologique de la région d'Andlau http://louise.michel.g.free.fr/upload/thouand_e-47c5d70-Bac-2021-m-tropole-septembre9-21svtj1me3-exo2-datation-Alsace-.pdf |
||
|
À faire
pour le
25-11-2023
|
Bac blanc de 8 h 30 à 12 h
|
||
|
Commentaires
|
|||
|
Date
|
Jeudi 23 novembre 2023 |
Matière
|
SVT_THOUAND 136
|
|
Chapitre
|
2.2 Les traces du passé mouvementé de la Terre
|
Séquence
|
|
|
Activités
|

| ||
|
Résumé
|
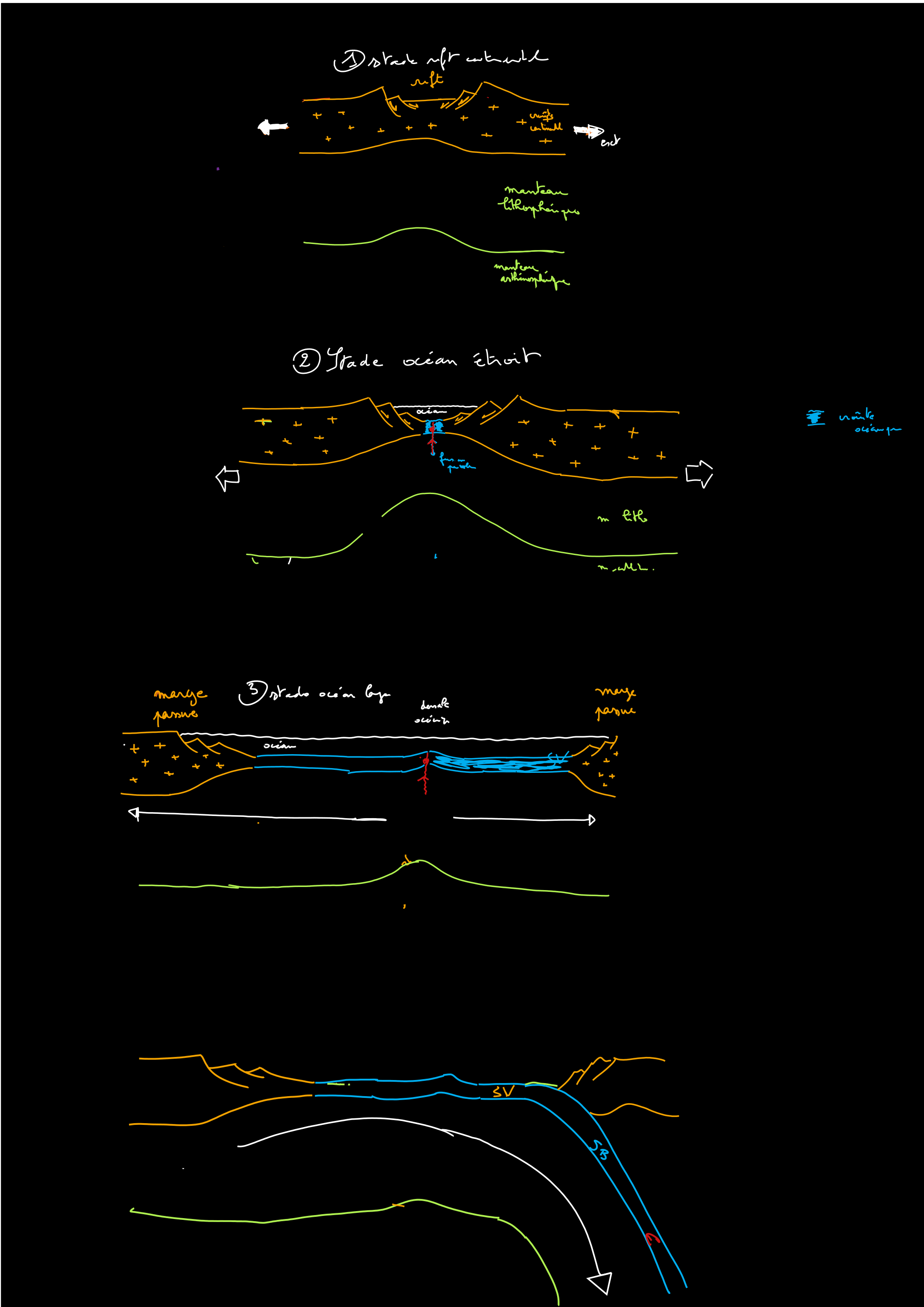
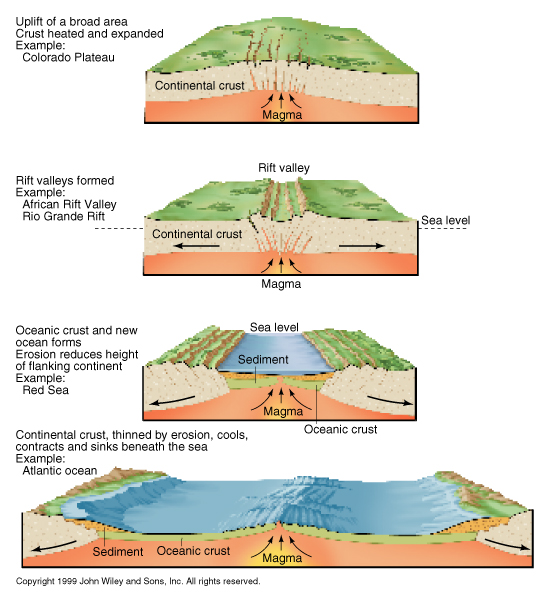
Cours III La fragmentation continentale et l’ouverture océanique [suite]
Si la distension continue, la lithosphère continentale devient tellement mince qu’elle se retrouve sous le niveau des océans : elle est alors immergée. Cet amincissement extrême provoque la mise en place d’un magmatisme basaltique à l’origine de la production de croûte océanique (stade océan étroit).Si la distension continue encore, le domaine océanique s’élargit, avec en son centre une dorsale séparant deux plaques divergentes (stade océan). Les marges passives de chaque côté de cet océan portent des marques des mouvements en extension (failles normales et blocs basculés) qui ont eu lieu lors de la fragmentation continentale (stade rift). La fragmentation continentale précède l’accrétion océanique (accrétion océanique = formation de lithosphère océanique par ajout progressif de matériel au niveau d’une dorsale). TD L’exercice 1 de l’épreuve écrite du bac Veillez à avoir un discours argumenté (expériences, observations,…) |
||
|
À voir
|
|
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 22 novembre 2023 |
Matière
|
SVT_THOUAND 135
|
|
Chapitre
|
2.2 Les traces du passé mouvementé de la Terre
|
Séquence
|
|
|
Activités
|
| ||
|
Résumé
|
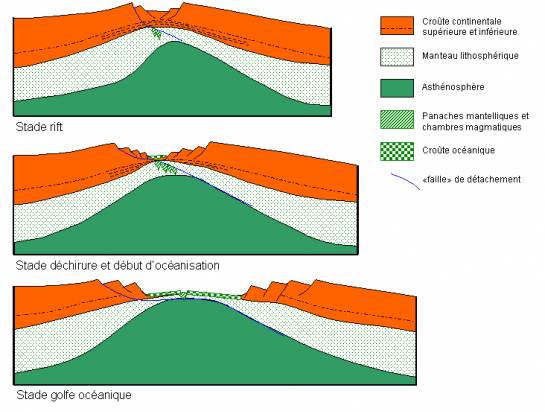
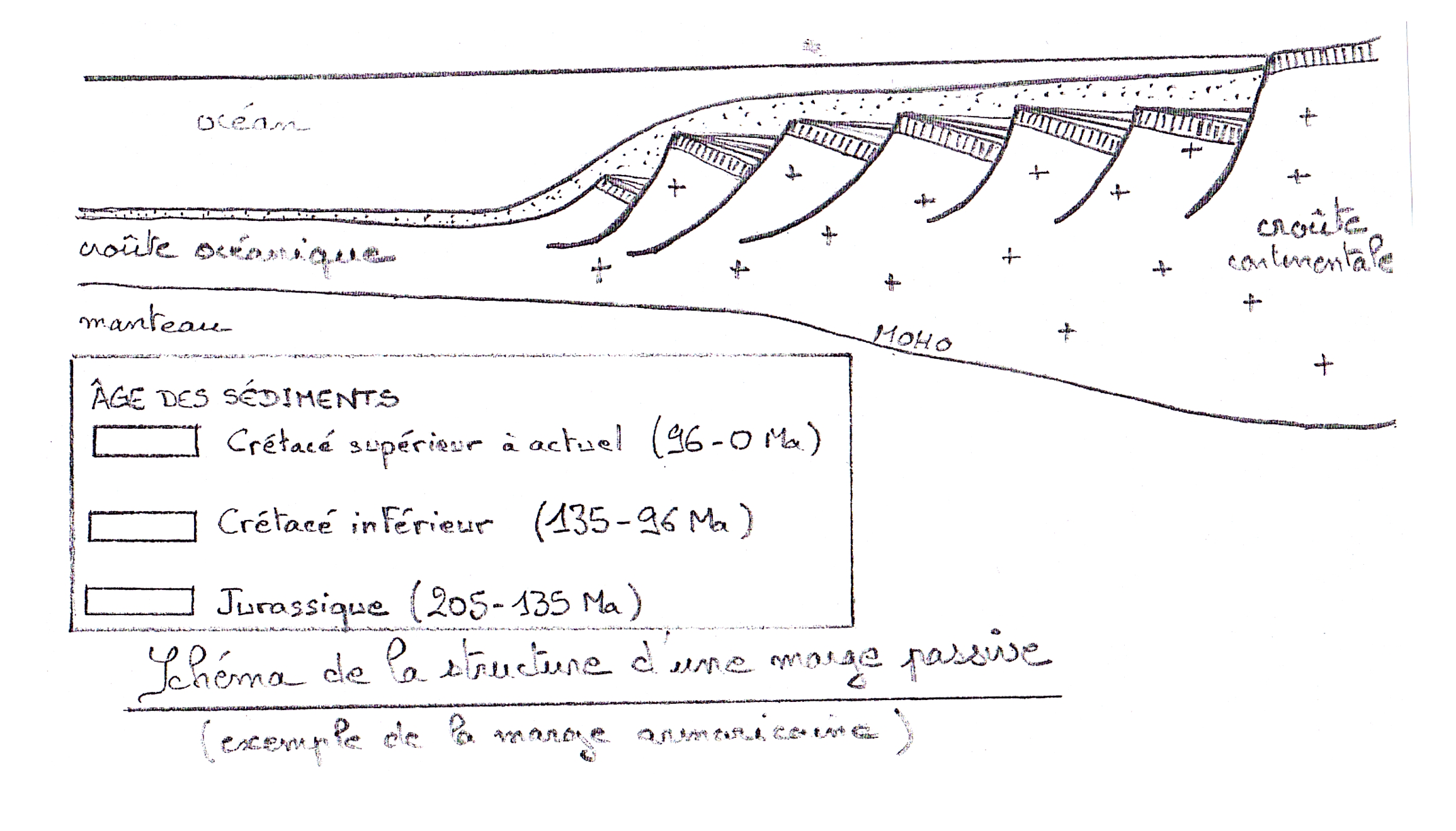
Cours III La fragmentation continentale et l’ouverture océanique [suite]
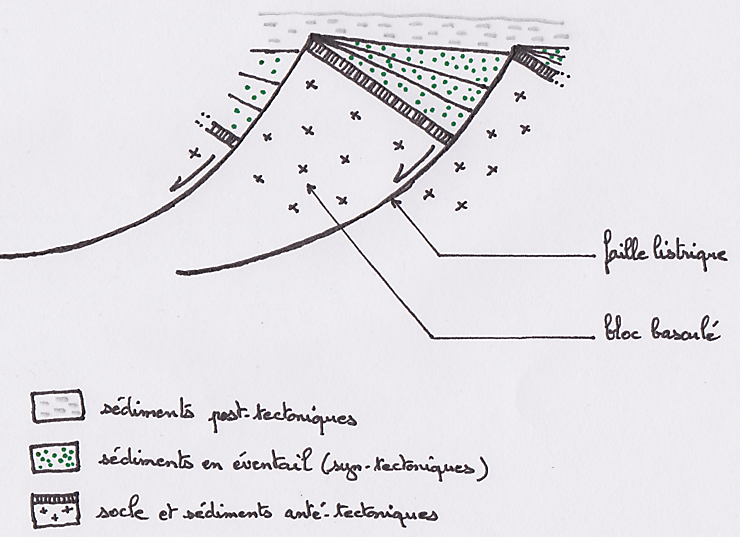
Schéma de la structure d’une marge continentale passive (exemple de la marge armoricaine)Au niveau d’une marge continentale passive, on observe : — des failles listriques : grandes failles normales recoupant la croûte sur une grande épaisseur, tendant à devenir horizontales en profondeur ; ne jouent plus actuellement (pas de séismes dans les marges passives) — des blocs basculés : grands blocs de croûte continentale, séparés par des failles listriques, qui se sont effondrés du côté du continent — des sédiments en éventail : recouvrent les blocs basculés. Leur disposition en éventail, c’est-à-dire avec une épaisseur plus importante d’un côté (vers le continent), indique qu’ils se sont déposés pendant que les blocs basculaient = sédiments syn-tectoniques. Les sédiments les plus anciens sont continentaux, les sédiments plus récents océaniques. Ces structures indiquent que la croûte continentale a subi des mouvements d’extension (= distension). La symétrie des marges passives de part et d’autre d’un océan, et le fait la croûte océanique les séparant soit postérieure au basculement des blocs indique que l’ouverture d’un océan est précédée par une phase de fragmentation continentale : des contraintes extensives provoquent un amincissement de la croûte continentale avec la formation de failles listriques et de blocs basculés symétriques par rapport à une vallée d’effondrement axiale = rift continental |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 17 novembre 2023 |
Matière
|
SVT_THOUAND 116
|
|
Chapitre
|
2.2 Les traces du passé mouvementé de la Terre
|
Séquence
|
|
|
Résumé
|
TD Sujet d’exercices 2 : Momies égyptiennes, isotopes et précipitations en Afrique intertropicale |
||
|
À voir
|
|||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Jeudi 16 novembre 2023 |
Matière
|
SVT_THOUAND 124
|
|
Chapitre
|
2.2 Les traces du passé mouvementé de la Terre
|
Séquence
|
|
|
Résumé
|
Cours II La recherche de lithosphère océanique disparue
Complexes ophiolitiques :
Un complexe ophiolitique est une formation formée d’une succession des roches suivantes (du sommet vers la base) :— basalte en coussins (= pillow-lavas) — basalte en filons — gabbros, métamorphisés (= métagabbros), de faciès schiste vert — péridotites serpentininisées (= métamorphisées, de faciès schiste vert) = serpentine Les complexes ophiolitiques peuvent ainsi être interprétés comme des lambeaux de lithophère océanique. On les observe notamment dans la zone centrale des chaînes de montagnes de collision, à la suture entre les deux plaques lithosphériques. Ceci suggère l’ existence passée d’une lithosphère océanique entre les deux domaines continentaux entrés en collision. Exemple : Complexe ophiolitique du Mont Chenaillet, dans les Alpes. Datation par radiochronologie des gabbros : - 160 Ma -> il existait à cette époque un océan entre les plaques eurasiatiques et africaines (Italie) L’émergence d’ophiolites peut résulter du phénomène d’ obduction : le lambeau de lithosphère océanique chevauche la lithosphère continentale sans avoir subduit. Dans ce cas, les ophiolites présentent un métamorphisme de faciès schiste vert (exemple de l’ophiolite du Chenaillet). Par ailleurs, les ophiolites peuvent avoir été exhumées après avoir entamé une subduction, qui se traduit par un métamorphisme de faciès schiste bleu / éclogite. La lithosphère océanique pincée entre les deux lithosphère continentales peut alors être exhumée. Rappels sur le métamorphisme (notamment celui lié à la subduction)
Roche métamorphique = issue de la transformation (à l’état solide : pas de fusion) d’une autre roche. Cette transformation se fait principalement sous l’effet de la pression (directement proportionnelle à la profondeur) et de la température (qui augmente généralement avec la profondeur, mais selon un gradient variable et non linéaire).Le métamorphisme peut se traduire par une modification de la texture de la roche, avec par exemple la mise en place d’une schistosité : les minéraux s’orientent dans des plans parallèles, ce qui forme des feuillets de minéraux. Il se traduit également par des réactions chimiques entre minéraux (lentes car à l’état solide). Ces réactions ont lieu dans des domaines précis de pression et de température : les faciès métamorphiques (associations de minéraux métamorphiques caractéristiques d’un domaine de pression-température particulier) permettent ainsi de reconstituer l’histoire de la roche (par quels domaines de pression-température elle est passée). N.B. : les réactions sont lentes, certains minéraux peuvent ainsi ne pas avoir été transformés : minéraux reliques. Les minéraux néoformés forment alors des auréoles métamorphiques autour des minéraux reliques. Au cours de son vieillissement, la lithosphère subit un métamorphisme lié à son refroidissement et son hydratation : métamorphisme de faciès schiste vert Au cours de sa subduction, elle subit un métamorphisme lié à l’augmentation de pression : métamorphisme de faciès schiste bleu, puis éclogite à plus grande profondeur III La fragmentation continentale et l’ouverture océanique
marge continentale : limite entre lithosphère continentale et lithosphère océaniquemarge continentale passive : marge continentale située à l’intérieur d’une plaque lithosphérique (=> pas d’activité sismique, ni volcanique ; par opposition avec une marge active, où la subduction est à l’origine de séismes et de magmatisme) plateforme continentale : zone formée de croûte continentale immergée, c’est-à-dire sous le niveau des océans. Attention, la marge continentale peut être située assez loin du littoral. |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 15 novembre 2023 |
Matière
|
SVT_THOUAND 134
|
|
Chapitre
|
2.2 Les traces du passé mouvementé de la Terre
|
Séquence
|
|
|
Résumé
|
Cours Chapitre 2 — Les traces du passé mouvementé de la Terre
I Des domaines continentaux présentant des âges variés
La croûte continentale présente des domaines d’âges variés, parfois très anciens (jusqu’à ~ 4 Ga). La présence de roches très anciennes, contrairement à la croûte océanique, suggère qu’il n’existe pas (ou peu) de recyclage de croûte continentale (alors que la croûte océanique âgée finit toujours par entrer en subduction).La croûte continentale est principalement constitué d’un socle : ensemble de roches métamorphiques (gneiss, etc.) et magmatiques plutoniques (granite, etc.), sur 30 km d’épaisseur. Ces roches se sont formées dans la racine crustale d’anciennes chaînes de montagnes : ce sont des reliquats d’anciennes ceintures orogéniques. L’étude de la croûte continentale montre l’existence de cycles orogéniques successifs (orogenèse = formation d’une chaîne de montagnes). L’érosion de ces anciennes chaînes de montagnes a exhumé ces roches formées en profondeur. Ce socle a pu être ultérieurement recouvert de roches sédimentaires, pouvant former de vastes bassins sédimentaires (dont l’épaisseur ne dépasse pas 3 km : en volume total, les roches sédimentaires sont négligeables par rapport aux granitoïdes du socle). En France métropolitaine, le socle correspond à la racine crustale de la chaîne hercynienne = varisque, vaste chaîne de montagnes de collision mise en place au Carbonifère (- 350 à - 300 Ma) en lien avec la formation de la Pangée. Cette ceinture orogénique se retrouve également en Amérique du Nord et jusqu’un Asie orientale. Ce socle affleure dans certaines régions (Massif central, Massif armoricain) ; dans le Bassin parisien ou le Bassin aquitain, il est recouvert de roches sédimentaires. |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 10 novembre 2023 |
Matière
|
SVT_THOUAND 104
|
|
Chapitre
|
2.1 La datation – Le temps et les roches
|
Séquence
|
|
|
Problèmes
|
|
||
|
Résumé
|
TP TP Datation absolue de granites par radiochronologie Utilisation d’un tableur pour tracer la droite isochrone, obtenir son coefficient directeur et calculer l’âge de la roche Document de travail : http://louise.michel.g.free.fr/upload/thouand_e-462a968-Datation-granite-p-162-.ods TD Sujet d’exercice 2 : datation de projections de ponces de la caldéra de Haute-Dordogne |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Jeudi 09 novembre 2023 |
Matière
|
SVT_THOUAND 106
|
|
Chapitre
|
2.1 La datation – Le temps et les roches
|
Séquence
|
|
|
Résumé
|
Cours —II La chronologie absolue
Datation absolue = détermination de l’âge d’une rochePossible que depuis le début du XXe siècle (après la découverte de la radioactivité) ——II.1 Principe général de la radiochronologie
Les isotopes radioactifs se désintègrent régulièrement au cours du temps : élément père -> élément fils.La quantité d'élément fils formé au cours du temps suit une loi exponentielle, caractérisée par : – la demi-vie T (temps au bout duquel la moitié des éléments père se sont désintégrés) – la constante de désintégration du couple père/fils ( [latex]\mathrm{\lambda}[/latex], en [latex]\mathrm{an^{-1}}[/latex]) Lors de la solidification d’un magma, des éléments sont intégrés dans les minéraux et n’en sortent plus : à partir de ce moment, le système est fermé. La mesure de la quantité d’éléments père restants (ou d’éléments fils formés) permet ainsi de déterminer la date de la formation d’une roche magmatique. N.B. Seules les roches magmatiques peuvent être datées par radiochronologie (les roches sédimentaires et métamorphiques ne constituent pas des systèmes fermés). Un couple élément père / élément fils est appelé géochronomètre. ——II.2 Application pratique (ex. du couple rubidium/strontium)
Étude d’un géochronomètre particulier : le couple [latex]\mathrm{^{87}Rb/^{87}Sr}[/latex]– T = 42,2 Ga – [latex]\mathrm{\lambda=1,4.10^{-11}an^{-1}}[/latex] N.B. : on ne mesure pas la quantité absolue de [latex]\mathrm{^{87}Rb}[/latex] et de [latex]\mathrm{^{87}Sr}[/latex], mais leur quantité relative par rapport à un autre élément, le [latex]\mathrm{^{86}Sr}[/latex] dont la quantité est constante au cours du temps => ce sont des rapports isotopiques qui sont mesurés : [latex]\mathrm{\frac{^{87}Rb}{^{86}Sr}}[/latex] et [latex]\mathrm{\frac{^{87}Sr}{^{86}Sr}}[/latex] Problème : on ne connaît pas les quantités initiales de [latex]\mathrm{^{87}Rb}[/latex] et de [latex]\mathrm{^{87}Sr}[/latex] (de plus, tous les minéraux n'incorporent pas la même quantité de 87Rb). Leur quantité dans la roche ne peut donc pas directement permettre de déterminer l’âge de la roche. Résolution du problème : On utilise un graphique avec la quantité de [latex]\mathrm{^{87}Sr}[/latex] (élément fils) en ordonnées, et la quantité de [latex]\mathrm{^{87}Rb}[/latex] (élément père) en abcisses. On place sur ce graphique les points correspondant aux rapports isotopiques mesurés dans plusieurs minéraux de la roche à dater. Lors de la formation de la roche (t=0), tous les minéraux contenaient autant de [latex]\mathrm{^{87}Sr}[/latex] => sur le graphique, les points étaient alignés suivant une droite horizontale. Au cours du temps, dans chaque minéral la quantité de [latex]\mathrm{^{87}Rb}[/latex] diminue tandis que la quantité de [latex]\mathrm{^{87}Sr}[/latex] augmente => les points sont toujours alignés, mais la droite se « verticalise » (son coefficient directeur augmente). Plus le temps passe, plus le coefficient directeur de cette droite augmente. Déterminer ce coefficient directeur permet donc de déterminer le temps écoulé depuis la formation de la roche. Si l’on mesure les rapports isotopiques dans différents minéraux d’une même roche, et que l’on place les valeurs sur un graphique [latex]\mathrm{^{87}Sr = f(^{87}Rb)}[/latex], on obtient des points alignés selon une droite appelée droite isochrone. Du fait de la désintégration des éléments père en éléments fils, ces points restent alignés au cours du temps, et la droite isochrone se verticalise au fur et à mesure que le temps passe c’est-à-dire que son coefficient directeur augmente (à t=0, lorsque le système se ferme, la droite est horizontale, car tous les minéraux ont le même rapport isotopique pour le [latex]\mathrm{^{87}Sr}[/latex]). Le coefficient directeur a de la droite isochrone est lié au temps t écoulé depuis la fermeture du système par la relation :[latex]\mathrm{\boldsymbol{{t=\frac{ln(a+1)}{\lambda}}}}[/latex] où [latex]\mathrm{t}[/latex] est le temps en années, et [latex]\mathrm{\lambda}[/latex] la constante de désintégration du géochronomètre étudié en [latex]\mathrm{an^{-1}}[/latex] |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 08 novembre 2023 |
Matière
|
SVT_THOUAND 105
|
|
Chapitre
|
2.1 La datation – Le temps et les roches
|
Séquence
|
|
|
Problèmes
|
|
||
|
Résumé
|
Cours I La chronologie relative [suite]
I.2 Biostratigraphie
Les espèces n’existent que pendant certaine période de temps => les fossiles d’une roche sédimentaire peuvent être utilisés pour dater le dépôt des sédimentsCf. manuel pp. 126-127 Fossile stratigraphique = espèce disparue qui : — présentait une grande extension géographique (=> fossiles susceptibles d’être trouvés en de multiples endroits) — appartient à un groupe à évolution rapide (=> courte durée d’existence de l’espèce => caractéristique d’une période donnée) — présente de nombreux fossiles (si les fossiles sont très rares, ils ne peuvent être utilisés pour dater !) Principe de continuité : une strate présente le même âge sur toute son étendue => associations de fossiles stratigraphiques : utilisées pour caractériser des intervalles de temps Si une même association fossile est trouvée dans des formations éloignées géographiquement, on en déduit l’existence d’une corrélation temporelle entre ces formations. I.3 L’échelle stratigraphique
Paléontologie + biostratigraphie => établissement de l’ échelle stratigraphique = « échelle des temps géologiques » (cf. rabat de la première page du manuel)Dans l’échelle stratigraphique, les intervalles de temps sont délimités par des coupures établies sur des critères paléontologiques (apparition/disparition de groupes fossiles) Différents ordres de coupure : ères > périodes (= systèmes) > étages (= séries) 3 ères : — cénozoïque = « ère tertiaire » — mésozoïque = « ère secondaire » – paléozoïque = « ère primaire » Les limites entre les ères correspondent à des crises biologiques majeures Quelques repères temporels à avoir
-0,3 Ma = -300 000 ans : plus anciens Homo sapiens connus-7 Ma : séparation de la lignée humaine et de la lignée du chimpanzé -65 Ma : crise crétacé/tertiaire (K/T) = 5ème crise biologique majeure, disparition des dinosaures non aviens, des ammonites, etc. -355 à -300 Ma : Carbonifère (formation de la Pangée, orogenèse hercynienne, climat froid, dépôt de charbon) -540 Ma : début du Paléozoïque (Cambrien) = "ère primaire" (auparavant peu d’êtres vivants pouvant être fossilisés) -4 Ga : apparition de la vie sur Terre -4,55 Ga : formation de la Terre |
||
|
À voir
|
|
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 20 octobre 2023 |
Matière
|
SVT_THOUAND 98
|
|
Chapitre
|
1.2 La complexification des génomes : transferts horizontaux et endosymbiose
|
Séquence
|
|
|
Résumé
|
Évaluation Test sur l’alphabet grec TD Présentation de l’exercice 2 de l’épreuve écrite du baccalauréat
Exercice type bac
« Exercice 2 : La féminisation des populations de cloporte »http://louise.michel.g.free.fr/upload/thouand_e-4496295-Wolbachia-et-cloportes-2021-.pdf |
||
|
À voir
|
|||
|
À faire
pour le
08-11-2023
|
Faire le sujet Liban 2016 du polycopié « Florilège de sujets d’exercice 1 sur le chapitre 1.1 » (cf. lien ci-dessus)
Sur ce florilège, choisir un sujet de 2021, 2022 ou 2023. Réalisez uniquement les schémas attendus. [à faire sur une autre feuille ; pensez à bien indiquer le sujet choisi !] S’inscrire sur agora (par exemple ici : http://louise.michel.g.free.fr/test.html ou via le lien sur la page d’accueil), rejoindre le canal dédié au groupe de terminale spécialité SVT. Y poster vos schémas du sujet au choix, (en indiquant le sujet). |
||
|
Commentaires
|
|||
|
Date
|
Jeudi 19 octobre 2023 |
Matière
|
SVT_THOUAND 99
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
TD Correction des exercices 2, 3, 7 pages 51-53 Correction du TP Brassage intrachromosomique chez la drosophile (6 oct.) Orientation Présentation de la CPGE BCPST (classe prépa. « agro ») et des grandes écoles auxquelles elle mène |
||
|
À voir
|
Prépa BCPST : présentation sur le site de l’ONISEP |
||
|
À faire
pour le
26-10-2023
|
Pour les élèves qui n’ont pas eu la moyenne à l’évaluation sur la méiose (du 6 oct.), venir demain refaire les schémas d’une méiose avec deux gènes liés
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 18 octobre 2023 |
Matière
|
SVT_THOUAND 100
|
|
Chapitre
|
2.1 La datation – Le temps et les roches
|
Séquence
|
|
|
Résumé
|
Cours Thème 2 — À la recherche de passé géologique de notre planète
Chapitre 1 — La datation - Le temps et les roches
Chronologie relative/absolue– chronologie relative : datation des événements/structures les uns par rapport aux autres (plus récent / plus âgé, sans avoir connaissance de leur âge – chronologie absolue : attribution d’un âge précis Jusqu’au début du XXe siècle (suite à la découverte de la radioactivité), seules des datations relatives étaient possibles. Rappel sur les différents types de roches — roches magmatiques : issues de la solidification d’un magma (= roche en fusion) Roches magmatiques volcaniques : refroidissement rapide du magma (à la surface) => texture microlitique. Ex. : basalte, rhyolite Roches magmatiques plutoniques : refroidissement lent du magma (en profondeur, dans un pluton) => texture grenue. Ex. : gabbro, granite — roches sédimentaires : issues d’un sédiment (= dépôt) Forment des couches superposées : strates Ex. : calcaire, grès — roches métamorphiques : issues de la transformation, à l’état solide, d’une autre roche (magmatique, sédimentaire ou métamorphique). Transformations généralement dues à des augmentations de pression et/ou de température Ex. : schistes Érosion Les roches sont dégradées (mécaniquement et chimiquement), principalement sous l’action de l’eau ; les reliefs tendent ainsi à disparaître : c’est l’érosion. Les produits de cette altération sont transportés plus ou moins loin et finissent par sédimenter. I La chronologie relative
I.1 Relations géométriques
Principes fondamentaux de la datation relative :— principe de continuité : une strate présente le même âge sur toute son étendue — principe de superposition : Une couche sédimentaire (ou une coulée volcanique) est plus jeune que la couche qu’elle recouvre — principe de recoupement : Une structure géologique (faille, filon volcanique, etc.) est plus jeune que la structure qu’elle recoupe — principe d’ inclusion : Un objet géologique inclus dans un autre est plus ancien que la structure qui l’entoure Présentation du « défi de Lyell » : jeu en ligne pour comprendre les principes de la datation relative : https://www.pedagogie.ac-nice.fr/svt/productions/defi-lyell/index.htm Analyse d’exemples RAPPEL : FAILLES NORMALES ET INVERSES
Description de ces deux types de failles, lien avec avec les contraintes qui en sont à l’origine :— contraintes en extension => faille(s) normale(s) => allongement des terrains — contraintes en compression => faille(s) inverses(s) => raccourcissement des terrains Pour les reconnaître, retrouvez les contraintes qui ont pu en être à l’origine, ou sachez qu’une faille normale est une faille où le bloc supérieur (celui au-dessus du plan de faille) a été animé d’un mouvement relatif vers le bas [vers le haut pour une faille inverse] |
||
|
À voir
|
Les différents types de roches, avec les informations que l’on peut en retirer Conférence (~ 1 h) sur la datation relative, par André Schaaf en 2004 [pour les plus passionné·e·s] Illustrations des principes de la datation relative : des cas concrets
[failles] |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 13 octobre 2023 |
Matière
|
SVT_THOUAND 93
|
|
Chapitre
|
1.2 La complexification des génomes : transferts horizontaux et endosymbiose
|
Séquence
|
|
|
Résumé
|
Cours II Origine endosymbiotique des mitochondries et plastes [suite]
Les organites sont transmis de génération en génération, par les gamètes femelles uniquement : hérédité cytoplasmique (pas de brassage génétique, au contraire du génome nucléaire !). TD Analyse des articles de la revue de presse
Articles en lien à la séance du 12 octobre TD Complexification des génomes
Correction des exercices 1, 2, 3 |
||
|
À voir
|
Endosymbiose, hérédité cytoplasmique : |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Jeudi 12 octobre 2023 |
Matière
|
SVT_THOUAND 85
|
|
Chapitre
|
1.2 La complexification des génomes : transferts horizontaux et endosymbiose
|
Séquence
|
|
|
Résumé
|
Cours I.3.c Applications biotechnologiques [suite]
— OGM pour l’industrie (agro-alimentaire). Ex. p. 64 : des transferts de gènes pour l’industrie laitière— thérapie génique II Origine endosymbiotique des mitochondries et plastes
symbiose = association étroite entre deux êtres vivants, à bénéfice mutuelendosymbiose = symbiose où l’un des symbiotes est situé à l’intérieur des cellules de l’autre symbiote Observations au niveau des mitochondries et chloroplastes (cf. pp. 62-63) : – présence d’ADN et de ribosomes, de type bactérien – division autonome par scissiparité (type de mitose caractéristique des bactéries) – présence de deux membranes, la membrane interne de type bactérien — génome proche de celui de certaines eubactéries, mais très éloigné du génome nucléaire de la cellule hôte (p. 63 doc. 5) => hypothèse d’une origine endosymbiotique (endocytose d’une bactérie par une cellule eucaryote) de ces organites, à partir de bactéries libres qui ont été intégrées dans le cytoplasme de cellules eucaryotes Schéma du mécanisme à l’origine de l’endosymbiose Il s’agit d’une symbiose entre cellule eucaryote et bactérie, car : — la bactérie apporte à la cellule eucaryote des capacités biochimiques particulières (respiration cellulaire pour la mitochondrie ; photosynthèse pour les chloroplastes) — la cellule eucaryote apporte à la bactérie la fourniture de nutriments, et la protection Les organismes eucaryotes ont ainsi un génome de deux origines : un génome nucléaire d’origine eucaryote, et un génome mitochondrial d’origine bactérienne (voire un troisième génome si présence de plaste) = génome chimérique Après l’endocytose initiale, les bactéries endosymbiotiques ont été transmises de génération en génération, par voie cytoplasmique. p. 63 doc. 6,7 : Au fil des générations, il y a eu régression progressive du génome de l’organite, notamment suite à un transfert dans le génome nucléaire (et pertes de gènes faisant doublon avec ceux du génome nucléaire) Les organites sont transmis de génération en génération, par les gamètes femelles uniquement : hérédité cytoplasmique (pas de brassage génétique, au contraire du génome nucléaire !). |
||
|
À voir
|
Thérapie génique : |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 11 octobre 2023 |
Matière
|
SVT_THOUAND 86
|
|
Chapitre
|
1.2 La complexification des génomes : transferts horizontaux et endosymbiose
|
Séquence
|
|
|
Résumé
|
Cours I.3.b Mécanisme de diversification du vivant [suite]
Exemple :p. 60 doc. 1 à 5 Chez l'humain et de nombreux mammifères (et le lézard mabuya), le gène de la syncytine (protéine permettant la formation du placenta par fusion de nombreuses cellules) est d'origine virale. Dans le cas des primates, le gène de la syncytine a été acquis par transfert horizontal d’un gène du virus MPMV codant pour la protéine d’enveloppe virale (qui permet la fusion de cette enveloppe avec la membrane plasmique de la cellule infectée). Ce transfert horizontal du gène viral de la syncytine a eu lieu de nombreuses fois chez les mammifères (p. 60 doc. 5). => les transferts horizontaux de gènes sont assez courants au cours de l’évolution Chez l'humain, 8 % du patrimoine génétique est d'origine virale ! => les transferts horizontaux ont un rôle important dans l’histoire évolutive, en permettant l’apparition de nouveaux caractères fondamentaux (sans transfert horizontal, il n’aurait pu y avoir apparition du placenta chez les mammifères) I.3.c Applications biotechnologiques
— OGM agricoles (transfert dans une plante cultivée d’un gène de production d’un insecticide, ou d’un gène de résistance à un herbicide).Exemple : maïs Bt, à qui l’humain a transféré un gène issu de la bactérie Bacillus thurigiensis, codant pour une protéine insecticide. (histoire des OGM ; glyphosate et Monsanto) — OGM à destination thérapeutique Exemple de la production d’insuline humaine (nécessaire pour les diabétiques de type 1), par transfert du gène de l’insuline humaine dans des micro-organismes faciles à cultiver en masse |
||
|
À voir
|
Transferts horizontaux :
Transferts horizontaux :
Transferts horizontaux :
Transferts horizontaux : |
||
|
À faire
pour le
12-10-2023
|
Exercices 2 et 3 du TD « Complexification des génomes »
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 06 octobre 2023 |
Matière
|
SVT_THOUAND 80
|
|
Chapitre
|
1.1 / 1.2
|
Séquence
|
|
|
Résumé
|
Évaluation Test de connaissances Schémas d’enjambement inégal Gamètes produits par un individu hétérozygote pour deux gènes liés Schémas d’une méiose avec enjambement entre deux gènes liés TP TP Brassage intrachromosomique chez la drosophile Répondre par écrit à la première étape : Élaborer une stratégie de résolution afin de vérifier si la fréquence des enjambements dépend de la distance entre les gènes, en proposant quels croisements vous compteriez réaliser, en dressant les tableaux de fécondation correspondant, et en précisant les résultats attendus. TD L’origine endosymbiotique des chloroplastes et mitochondries À partir des documents pp. 62-63, listez les arguments qui suggèrent que les chloroplastes et les mitochondries ont pour origine des bactéries, qui se trouvent dorénavant dans les cellules eucaryotes. |
||
|
À voir
|
|||
|
À faire
pour le
11-10-2023
|
— revoir l’alphabet grec
— réaliser l’exercice 1 du TD « Complexification des génomes » (cf. énoncé ci-dessus) — lire au moins deux articles parmi ceux placés dans la rubrique « À voir » des séances précédentes (ou parmi ceux de la revue de presse concernant les chapitres 1.1 et 1.2). Préparer des notes en vue d’une courte présentation orale |
||
|
Commentaires
|
|||
|
Date
|
Jeudi 05 octobre 2023 |
Matière
|
SVT_THOUAND 78
|
|
Chapitre
|
1.2 La complexification des génomes : transferts horizontaux et endosymbiose
|
Séquence
|
|
|
Résumé
|
Cours I.2 Processus aboutissant à des transferts horizontaux [suite]
I.2.a Chez les bactéries [suite]
I.2.a. [latex]\mathrm{\gamma}[/latex] Transfert viral Description du mécanisme : cf. I.2.b (transferts horizontaux chez les eucaryotes) I.2.b Chez les eucaryotes
Processus principal : transfert viralRappel sur les virus Schématisation des différents mécanismes de transfert viral : — insertion d’un gène viral dans le génome de la cellule (conservé après l’infection, à condition que la cellule infectée n’ait pas été détruite) — virus servant de vecteur de gène entre deux taxons différents (particule virale ayant intégré par erreur un gène de la cellule infectée, qui va ainsi pouvoir être transféré dans une autre cellule, possiblement d’une autre espèce, si cette particule virale l’infecte) I.3 Conséquences des transferts horizontaux
I.3.a Transfert de gènes de résistance à des antibiotiques entre bactéries
Remarque :Importance des transferts horizontaux chez les bactéries (qui ne se reproduisent pas de manière sexuée => pas de brassage) : sont une source de diversité génétique chez les bactéries Problème de santé publique : possibilité de transfert d’un gène de résistance aux antibiotiques depuis une bactérie non pathogène (caractère sélectionné par l’un usage excessif d’antibiotiques) vers une bactérie pathogène => apparition de souches de bactéries pathogènes résistantes aux antibiotiques I.3.b Mécanisme de diversification du vivant
= apparition de nouveaux caractères dans un taxonExemple : p. 60 doc. 1 à 5 : analyse de documents sur l’origine du gène de la syncytine |
||
|
À voir
|
|
||
|
À faire
pour le
06-10-2023
|
Préparer évaluation de connaissances :
— schémas de méiose pour deux gènes indépendants, ou deux gènes liés (avec ou sans enjambements : cf. correction du devoir du 15 septembre ci-dessus) — schéma d’enjambement inégal — schémas des mécanismes de transfert horizontal chez les bactéries — maîtriser les tableaux de fécondation d’un croisement-test (entre autres) |
||
|
Commentaires
|
|||

|
Date
|
Mercredi 04 octobre 2023 |
Matière
|
SVT_THOUAND 87
|
|
Chapitre
|
1.2 La complexification des génomes : transferts horizontaux et endosymbiose
|
Séquence
|
|
|
Résumé
|
Cours Chapitre 1.1 L’origine du génotype des individus [suite]
Conclusion
Lors de la reproduction sexuée, les brassages génétiques (lors de la méiose et lors de la fécondation) sont à l’origine de la diversité génétique des descendants, qui présentent tous une combinaison d’allèles unique.Ces brassages sont ainsi à l’origine de la diversité génétique intraspécifique. Les anomalies lors de la méiose peuvent également être la source de génotypes originaux, donc source de diversité génétique. Dans certains cas (cf. famille multigénique), ces anomalies lors de la méiose peuvent même être à l’origine de nouveaux gènes et donc de nouveaux caractères : elles sont alors source de diversification du vivant Thème 1, chapitre 2 — La complexification des génomes : transferts horizontaux et endosymbiose
I Transferts horizontaux
N.B. : Transfert vertical de gène = transfert de gène lors de la reproduction sexuée, de parent à descendant (cf. Chapitre 1.1).Les gènes d’un individu sont ainsi issu pour moitié de l’un de ses parents, pour moitié de l’autre parent ; et ainsi de suite (les gènes d’un parent proviennent de ses deux parents, etc. : ceci dure depuis environ 4 Ga…). Transfert horizontal de matériel génétique = échange de matériel génétique hors de la reproduction sexuée (entre deux individus qui peuvent être de taxons différents) N.B. taxon = groupe d’êtres vivants possédant des caractères communs issus d’un ancêtre commun (espèce, genre, famille, ordre, classe, etc.) I.1 Mise en évidence à partir d’arbres phylogénétiques
Arbre phylogénétique = arbre indiquant les degrés de ressemblances, et donc de parenté, entre taxons ou entre molécules (gènes/protéines)Exemples d’arbre phylogénétique : p. 60 doc. 5 : arbre phylogénétique indiquant les parentés entre différents taxons de vertébrés p. 60 doc. 7 : arbre phylogénétique indiquant les parentés entre différentes enzymes (= protéines catalysant des réactions biochimiques) Comparaison avec les langues Comme pour les gènes, les langues sont principalement transmises par transfert vertical : on peut ainsi remonter de génération en génération, et observer leur évolution, et parfois leur séparation en deux langues (cf. spéciation en biologie). On peut donc reconstituer un arbre phylogénétique des langues (cf. document ci-dessous). -> distinction entre phylogénie des langues et phylogénie d’un mot : si un mot est très proche dans deux langues très éloignées phylogénétiquement (ex. : toubib en arabe et en français), ceci indique qu’il ne provient pas de la langue ancestrale commune (pas de transmission verticale), mais qu’il est passé d’une langue à l’autre (transmission horizontale). Mise en évidence de transferts horizontaux de gènes p. 69 ex. 2 Observation : certains nématodes possèdent des enzymes (polygalacturonases) capables de digérer la paroi des cellules végétales ; les gènes codant pour ces enzymes chez ces nématodes sont très proches des gènes de bactéries (doc. 4), alors que les nématodes sont des eucaryotes, dont le génome est très éloigné de celui des bactéries. Si ce gène avait été transmis verticalement, depuis le dernier ancêtre commun aux nématodes et aux bactéries, les gènes des nématodes et des bactéries seraient beaucoup plus différents (de plus, tous les nématodes posséderaient ce gène) Interprétation : chez ces nématodes, les gènes codant pour ces enzymes proviennent d'un transfert horizontal de gènes de bactéries. Ce transfert horizontal a eu lieu quatre fois chez les nématodes (doc. 3). I.2 Processus aboutissant à des transferts horizontaux
I.2.a Chez les bactéries
Cf. p. 58I.2.a. [latex]\mathrm{\alpha}[/latex] Transformation = intégration dans le génome bactérien d’ADN libre p.58 doc. 1,2 : expériences montrant la transformation chez des pneumocoques (expériences historiques : ce sont elles qui ont participé à démontrer que c’était l’ADN qui portait l’information génétique) I.2.a. [latex]\mathrm{\beta}[/latex] Conjugaison = transfert de plasmide entre deux bactéries, via un pont cytoplasmique (plasmide : molécule d’ADN en plus du chromosome bactérien, comportant des gènes non essentiels à la survie de la bactérie) |
||
|
À voir
|
|||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 29 septembre 2023 |
Matière
|
SVT_THOUAND 65
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
TD Analyses génétiques
Les maladies génétiques sont dues à une mutation sur un gène à l’origine d’un allèle morbide (= responsable de la maladie, par opposition à un allèle sain) Transmission des maladies génétiques
cf. p. 40 doc. 1Transmission récessive ou dominante — transmission dominante (= l’allèle morbide est dominant sur l’allèle sain) - un individu hétérozygote est malade - un individu malade a nécessairement un (au moins) parent malade - probabilité d’un individu malade (hétérozygote) d’avoir un descendant malade : 1/2 Ex. : maladie de Huntington (les allèles persistent dans la population, car la mortalité a lieu après 40 ans, donc après la reproduction) — transmission récessive (= l’allèle morbide est récessif) - seuls les individus homozygotes sont malades - un individu malade peut avoir deux parents sains, à condition qu’ils soient tous les deux hétérozygotes - probabilité d’un couple d’hétérozygotes (donc sains) d’avoir un enfant malade : malade : 1/4 [cf. tableau de fécondation] Transmission autosomique ou gonosomique — transmission gonosomique (= le gène est porté par un gonosome = chromosome sexuel) Gène sur le chromosome Y : Cas rare (peu de gènes sur le Y) ; n’affecte que les mâles Gène sur le chromosome X : Maladies plus fréquentes chez les mâles (un seul X, donc la possession d’un seul allèle morbide suffit pour être affecté), à qui l’allèle morbide a été transmis par la mère (chez un individu XY, le Y provient de son père, et le X de sa mère) Exemples : certains daltonismes (le gène des opsines sensibles au rouge, et celui des opsines sensibles au vert sont portés par le X, cf. cours sur les familles multigéniques) ; hémophilie — transmission autosomique (= le gène est porté par un autosome = chromosome non sexuel) Exercice d’application : la mucoviscidose
p. 40 doc. 3À partir de l’arbre généalogique, déterminer le type de transmission de la mucoviscidose, et déterminer la probabilité de l’enfant à naître (III.3) d’être malade. -> pratique du raisonnement par l’absurde. Exemple : si la transimission était dominante, alors un deux individus I.1 et I.2 serait atteint ; or ils sont sains ; l’hypothèse « transmission dominante » est donc invalidée. -> réalisation d’un tableau de fécondation (après avoir démontré que les parents de III.3 sont hétérozygotes, puisqu’ils sont sains mais ont déjà eu un enfant atteint) Remarque
Origine du caractère récessif ou dominant d’un allèle [non traité avec le groupe 2] |
||
|
À faire
pour le
04-10-2023
|
Réaliser (sur une feuille à part) les exercices 2, 3, 7 pages 51-53
|
||
|
Commentaires
|
|||
|
Date
|
Jeudi 28 septembre 2023 |
Matière
|
SVT_THOUAND 59
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
Cours III.2 Anomalie lors d’un enjambement [suite]
III.2.a Duplication d’un gène suite à en enjambement inégal
Schéma des conséquences d’un enjambement inégal en prophase I=> Un enjambement inégal peut aboutir à la duplication d’un gène dans un gamète (ou à la délétion d’un gène). III.2.b Conséquences des duplications d’un gène (non suivies de mutations)
La duplication d’un gène peut modifier le phénotype, de manière négative, voire létale.Exemple : la duplication du gène bar chez la drosophile entraîne une réduction drastique de la taille de l’œil. Mais il existe des cas ou la possession de plusieurs exemplaires identiques d’un gène (duplications non suivies de mutations) peut procurer un avantage reproductif à leur porteur, ce génotype étant alors sélectionné au fil des générations. Exemples : — moustiques : acquisition d’une résistance à certains insecticides par duplication du gène Est-2 (cf. p. 42 doc. 2,3) — éléphants : duplications du gène p53 => nombreux exemplaires du gène => plus faible probabilité pour une cellule tumorale => pas plus de cancers que les petits mammifères, malgré leur grand nombre de cellules — chien : nombreuses duplications du gène de l’amylase (jusqu’à 34 exemplaires, contre 2 chez les loups) codant pour une enzyme digérant l’amidon => meilleure capacité à digérer l’amidon présent dans les restes alimentaires des humains. NB : la même évolution a eu lieu chez… les humains (cf. http://louise.michel.g.free.fr/afficher_choix4.php?id=421) III.2.b Duplications de gène et famille multigénique
Suite à une duplication de gène, les deux exemplaires du gène sont identiques. Au fil des générations, chaque gène subit des mutations différentes. Ces mutations peuvent conduire l’ un des gènes à coder pour une protéine différente (on considère alors qu’il est devenu un gène différent), possédant une nouvelle fonction.famille multigénique = ensemble de gènes issus de duplications d’un gène ancestral, suivies de mutations Exemples : — les 3 gènes des pigments des cônes de la rétine (cf. p. 53 ex. 6) : proviennent de deux duplications d’un gène ancestral, suivies de mutations (et de translocation sur le gène X). La seconde duplication a eu lieu chez un primate ancestral, ce qui permet aux primates d’avoir un pigment absorbant dans le vert et un pigment absorbant dans le rouge => capacité à distinguer le rouge du vert — gènes de la vasotocine, l’ADH, l’ocytocine Conclusion sur les anomalies lors de la méiose
— souvent létales ou sources d’anomalies du phénotype des descendants (trisomie, monosomie par exemple)Mais : — peuvent au contraire être source d’un avantage reproductif (duplications du gène de résistance chez les moustiques, du gène p53 chez les éléphants, du gène de l’amylase chez le chien et l’humain par exemple) — peuvent être source de diversification du vivant (exemple de la duplication d’un gène suite à un enjambement inégal, suivie de mutations) diversification du vivant = apparition d’un nouveau caractère dans une espèce (cf. chapitres suivants). À ne pas confondre avec la diversité génétique (étudiée dans ce chapitre) = diversité des combinaisons d’allèles entre individus d’une même espèce (et donc diversité des phénotypes). |
||
|
À voir
|
|||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 27 septembre 2023 |
Matière
|
SVT_THOUAND 58
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
TP TP Brassage génétique chez la drosophile Correction Grand oral Possibilité de sujet sur l’histoire du chromosome Y, à l’évolution très rapide car n’est pas stabilisé par le brassage dû aux enjambements (il n’a jamais de chromosome homologue !). Par ailleurs, il est directement issu du père, sans brassage, ce qui permet de remonter au chromosome ancestral par voie patrilinéaire. Cf. articles de la revue de presse Cours III.1 Anomalie lors de la disjonction des chromosomes homologues (anaphase I) [suite]
Schéma des conséquences sur les gamètes d’une anomalie lors de la disjonction des chromosomes homologues en anaphase I [suite]=> gamètes avec un chromosome en deux exemplaires, gamètes avec un chromosome absent Après fécondation (avec un gamète normal) => production de zygotes présentant une trisomie ou une monosomie Remarque : une anomalie lors de la disjonction des chromatides en anaphase II produira les mêmes types de gamètes anormaux (mais seulement deux gamètes anormaux pour une méiose) Monosomie ou trisomie : généralement embryon non viable, sauf dans certains cas (trisomie pour le chromosome 21 par exemple ; monosomie ou trisomie pour les chromosomes sexuels : cf. « À voir » ci-dessous). Remarque : des anomalies lors de la mitose (non disjonction des chromatides d’un chromosome) peuvent conduire à un sous-clone présentant une trisomie ou une monosomie. Dans ce cas, toutes les cellules de l’individu ne possèdent pas le même caryotype : individu « mosaïque » (peut être à l’origine de certaines maladies). III.2 Anomalie lors d’un enjambement
III.2.a Duplication d’un gène suite à en enjambement inégal |
||
|
À voir
|
Exemples de monosomies/trisomies affectant les chromosomes sexuels |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 22 septembre 2023 |
Matière
|
SVT_THOUAND 51
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
TP Présentation détaillée de l’épreuve pratique du bac (ECE) Structure des deux types de sujet Description de la réflexion critique à apporter TP Brassage génétique chez la drosophile Utilisation de la loupe binoculaire Identifiez les quatre phénotypes sur les plaques avec les drosophiles Comptages avec Mesurim2 — Ouvrez Mesurim2 dans un navigateur web : https://www.pedagogie.ac-nice.fr/svt/productions/mesurim2/ — Dans la banque d’images, ouvrez l’image « Plaque de drosophiles (vg,b) F1BC #8 » (recherchez ces termes dans la barre de recherche) — Comptez les effectifs de chacun des quatre phénotypes — Indiquez vos résultats dans le tableau collaboratif : https://tab.louisematic.site/2022-2023_droso Sur votre copie : — dressez le tableau montrant le génotype (avec leurs proportions !) des gamètes produits par un individu ( vg+//vg eb+//eb ), puis de ceux d’un individu ( vg+,b+//vg,b ) [cette étape est facultative, mais elle est nécessaire si vous n’avez pas encore la maîtrise totale du cours — c’est en effet une pure question de cours : nous y avons vu quels sont les gamètes produits par un individu hétérozygote pour deux gènes indépendants, et pour deux gènes liés) — indiquez les tableaux de fécondation (avec les proportions !) attendus pour le croisement-test qui a été effectué, dans chacun des cas possibles : - si c’est le gène ebony qui est muté (indépendant du gène vg) : ( vg+//vg eb+//eb ) x ( vg//vg eb//eb ) - si c’est le gène black qui est muté (lié au gène vg*) : ( vg+,b+//vg,b ) x ( vg,b//vg,b ) — dressez le tableau des résultats de votre comptage (proportions des différents phénotypes parmi les descendants du croisement-test) — interprétez vos résultats en les comparant aux résultats attendus dans chacun des deux cas — discuter de la fiabilité des données recueillies |
||
|
À voir
|
Mesurim2 (lien pour lancer l’application dans un navigateur web) Tableau de résultats des comptages (à compléter par chacun·e) Sujet de TP (type ECE) « Brassage génétique chez la drosophile » |
||
|
À faire
pour le
27-09-2023
|
Terminer le TP (voir le résumé de la séance), et me le rendre (copie manuscrite + comptages sur le tableau collaboratif).
Seule la conclusion sera à faire ultérieurement, une fois que je vous aurai donné un document supplémentaire |
||
|
Commentaires
|
|||
|
Date
|
Jeudi 21 septembre 2023 |
Matière
|
SVT_THOUAND 46
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
Cours II.3 Le brassage génétique lors de la fécondation [suite]
[Partie à ne pas oublier dans le cas d’un sujet sur le brassage génétique ; au moins dresser un tableau de fécondation]fécondation = union de deux gamètes haploïdes, formant une cellule diploïde ( cellule-œuf = zygote) caryogamie = fusion des deux noyaux haploïdes formant un noyau diploïde Schéma des étapes de la fécondation (chez les animaux) rencontre des gamètes aléatoire => la fécondation amplifie le brassage génétique lors de la méiose Exemple Tableau de fécondation (= échiquier de croisement) pour le croisement ( A//a B//b ) × ( A//a B//b ) Chaque parent peut produire 4 gamètes différents (brassage génétique lors de la méiose), ce qui peut mener après fécondation à 9 génotypes différents au niveau des descendants. Cas de l’humain Deux humains, hétérozygotes pour leurs 20 000 gènes, peuvent produire [latex]\mathrm{2^{20000}\times2^{20000} = 2^{50000} = 1,6\times10^{12041}}[/latex] descendants différents possibles. [nombre très largement supérieur au nombre d’atomes dans l’univers] La probabilité que ces deux individus aient deux descendants de même génotype peut donc être considérée comme nulle. III Les anomalies lors de la méiose
III.1 Anomalie lors de la disjonction des chromosomes homologues (anaphase I)
ou lors de la disjonction des chromatides (anaphase II)
Schéma des conséquences sur les gamètes d’une anomalie lors de l’anaphase I=> gamètes avec un chromosome en deux exemplaires, et gamètes avec un chromosome absent TD Présentation des grandes lignes de l’épreuve pratique (ECE = évaluation des capacités expérimentales) |
||
|
À voir
|
Le nombre d’atomes dans l’univers : « seulement » [latex]10^{80}[/latex] ! |
||
|
À faire
pour le
22-09-2023
|
Maîtriser les brassages génétiques en vue d’une évaluation de TP
(+ alphabet grec) |
||
|
Commentaires
|
|||
|
Date
|
Mercredi 20 septembre 2023 |
Matière
|
SVT_THOUAND 47
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Activités
|
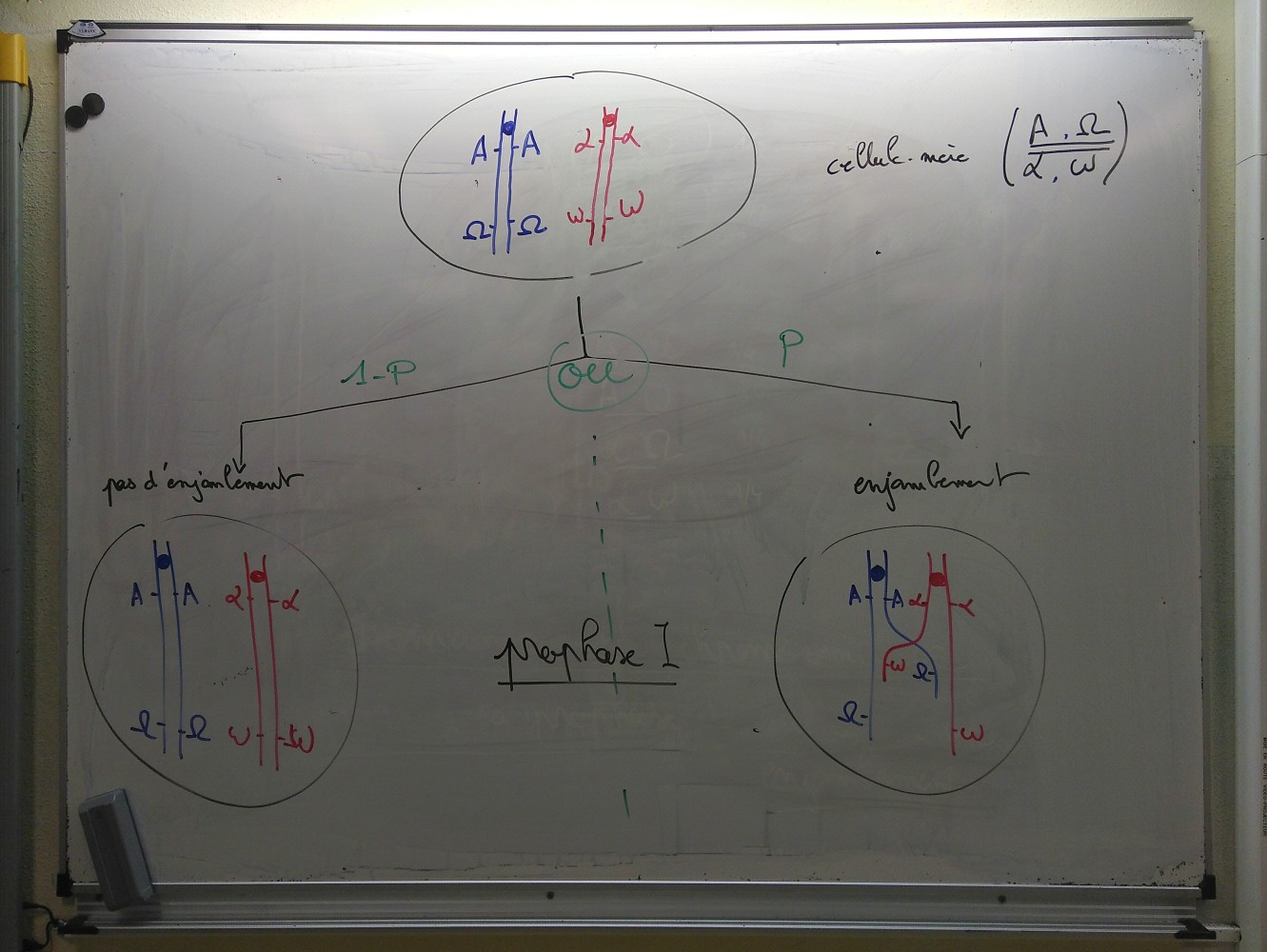

| ||
|
Résumé
|
Cours II.2.b Le brassage intrachromosomique [suite]
Schéma du remaniement chromosomique entraîné par un enjambementUn enjambement entraîne un remaniement chromosomique (= nouvelles combinaisons alléliques au sein d’un chromosome). Schéma du brassage intrachromosomique Tableau des gamètes produits par une cellule de génotype (A,B//a,b) : (A,B/) p>1/4 ; (a,b/) p>1/4 ; (A,b/) p<1/4 ; (a,B/) p<1/4 -> Le remaniement chromosomique provoqué par un enjambement lors de la prophase I conduit à la production de gamètes recombinés (présentant des combinaisons d’allèles absentes dans la cellule-mère). Ainsi, parmi les gamètes produits par un individu hétérozygote pour deux gènes liés, les 4 combinaisons alléliques possibles sont présentes, mais dans des proportions différentes : les deux gamètes parentaux sont plus abondants (p>1/4) que les gamètes recombinés (p<1/4) Remarque importante : Les enjambements sont très fréquents (nombreux chiasmas au sein d’un bivalent). Ce ne sont pas des anomalies de la méiose. (la probabilité qu’il y ait un enjambement entre deux gènes augmente avec la distance entre les gènes) II.2.c Conséquences des brassages génétiques lors de la méiose
Parmi les gamètes, toutes les combinaisons des allèles parentaux peuvent être présentes sont possibles.=> un individu hétérozygote pour N gènes peut produire [latex]\mathrm{2^N}[/latex] gamètes différents (possédant des combinaisons alléliques différentes) Exemple Individu hétérozygote pour 3 gènes (dont 2 liés) —> génotype des 8 gamètes différents qui peuvent être produits Cas de l’humain 20 000 gènes => un individu hétérozygote pour chacun des gènes peut produire [latex]\mathrm{2^{20000}}[/latex] gamètes différents (= [latex]\mathrm{4\times10^{6020}}[/latex], nombre très largement supérieur au nombre d’atomes dans l’univers). La probabilité que deux gamètes possèdent le même génotype peut donc être considérée comme nulle. II.3 Le brassage génétique lors de la fécondation
[Partie à ne pas oublier dans le cas d’un sujet sur le brassage génétique ; au moins dresser un tableau de fécondation]fécondation = union de deux gamètes haploïdes, formant une cellule diploïde ( cellule-œuf = zygote) |
||
|
À voir
|
Le nombre d’atomes dans l’univers : « seulement » [latex]10^{80}[/latex] ! |
||
|
À faire
|
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 15 septembre 2023 |
Matière
|
SVT_THOUAND 33
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
TD Réaliser tous les schémas d’une méiose pour une cellule de génotype ( [latex]\mathrm{A,\Omega//alpha,omega}[/latex]) [cellule hétérozygote pour deux gènes liés alpha et omega] En déduire quels sont (a priori — sans enjambement) les gamètes que peut produire cet individu. Cours II.2.a Le brassage interchromosomique [suite]
Tableau des gamètes produits par une cellule de génotype (A//a B//b) :(A/ B/) p=1/4 ; (a/ b/) p=1/4 ; (A/ b/) p=1/4 ; (a/ B/) p=1/4 => parmi les gamètes produits par un individu hétérozygote pour deux gènes indépendants (deux paires de chromosomes), les 4 combinaisons alléliques possibles sont présentes dans des proportions égales (1/4) (équiprobables) Conséquence du brassage interchromosomique : Un organisme possédant n paires de chromosomes peut produire [latex]\mathrm{\mathbf{2^n}}[/latex] gamètes différents ( [latex]\mathrm{2^n}[/latex] combinaisons de chromosomomes possibles).Un être humain peut ainsi produire [latex]\mathrm{\mathbf{2^{23} \simeq 8\times10^6}}[/latex] gamètes différents. II.2.b Le brassage intrachromosomique
Concerne des gènes liésLors de la prophase I, les chromosomes homologues sont étroitement appariés au sein d’un bivalent. Le croisement de leurs chromatides au niveau d’un chiasma peut conduire à un échange de portions de chromatide, appelé enjambement (= crossing-over). Évaluation Test sur l’alphabet grec |
||
|
À voir
|
|
||
|
À faire
pour le
20-09-2023
|
Réaliser tous les schémas, légendés, d’une méiose pour une cellule hétérozygote pour deux gènes indépendants (A//a B//b)
Vous pouvez vous inspirer des schémas ci-dessus (rubrique « À voir ») ou de vos schémas de première, en pensant bien à indiquer les allèles portés par chaque chromatide. Remarque : deux méioses différentes sont possibles, selon le sens de migration des différents chromosomes homologues. Vous ne représenterez qu’une méiose possible. |
||
|
Commentaires
|
|||

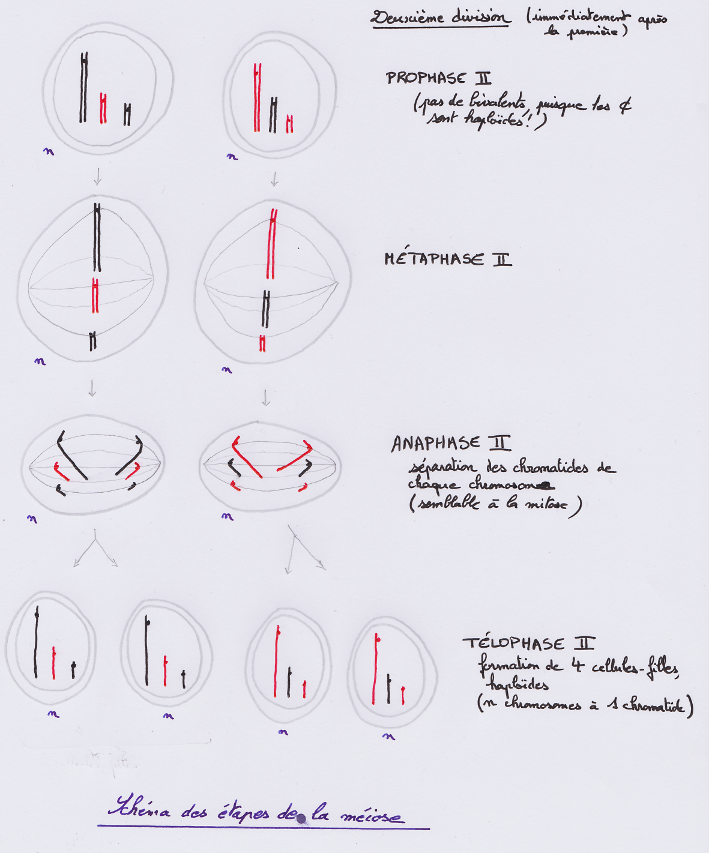
{kind=link}
|
Date
|
Jeudi 14 septembre 2023 |
Matière
|
SVT_THOUAND 26
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Activités
|


| ||
|
Résumé
|
Cours Annexe au chapitre 1.1 : notions fondamentales de génétique [suite]
— Vocabulaire de génétique [suite]- croisement-test = croisement entre une population (qui est « testée ») et une population d’individus tous homozygotes récessifs Dans le cas d’un croisement-test, le phénotype des descendants est déterminé par le génotype des gamètes de la population testée. Exemple : tableau de fécondation d’un croisement-test [latex]\mathrm{(x_1//x_2)\times(o//o)}[/latex] — méiose = processus qui forme, à partir d’une cellule-mère diploïde, quatre cellules-filles haploïdes ; se déroule dans les gonades, où elle forme les gamètes Précédée d’une réplication de l’ADN => au début de la méiose, les chromosomes sont doubles Deux divisions cellulaires successives : – première division (= division réductionnelle) : séparation des chromosomes homologues (-> forme 2 cellules haploïdes) – seconde division (= division équationnelle) : séparation des chromatides de chaque chromosome (similaire à la mitose) Chaque division cellulaire se déroule suivant les quatre phases successives : prophase, métaphase, anaphase, télophase Chapitre 1.1 [suite]
II.2 Les brassages génétiques lors de la méiose
À l’issue de la reproduction sexuée, chaque descendant possède une combinaison originale des allèles de ses parents : les allèles parentaux ont été mélangés = brassés.Les brassages génétiques lors de la méiose sont à l’origine de la diversité génétique des gamètes produits par un individu, c’est-à-dire de la diversité des combinaisons alléliques présentées par les gamètes. Un second brassage intervient ensuite, lors de la fécondation (cf. II.3), suite à la rencontre aléatoire des gamètes. La diversité génétique des descendants est donc due aux brassages lors de la méiose et au brassage lors de la fécondation. II.2.a Le brassage interchromosomique
Concerne des gènes indépendantsEst dû au fait que lors de l’ anaphase I, le sens de migration des chromosomes homologues est aléatoire et indépendant entre chaque paire. Schéma du brassage interchromosomique |
||
|
À faire
pour le
15-09-2023
|
Test sur tout l’alphabet grec
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 13 septembre 2023 |
Matière
|
SVT_THOUAND 16
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
Évaluation Test sur la méiose (avec une cellule hétérozygote pour un gène) Cours Annexe au chapitre 1.1 : notions fondamentales de génétique [suite]
— gènes indépendants (= sur des chromosomes différents) ou liés (= sur le même chromosome)— notation des allèles : plusieurs systèmes de notation des différents allèles d’un même gène Remarque : chez la drosophile, [latex]\mathrm{^+}[/latex] à côte du nom du gène désigne l’allèle sauvage (le plus courant dans les populations naturelles) ; l’allèle muté est désigné par l’absence de ce signe. Exemple pour le gène « ailes vestigiales » : l’allèle sauvage est noté [latex]\mathrm{vg^+}[/latex], l’allèle muté [latex]\mathrm{vg}[/latex]. — notation des phénotypes : toujours entre crochets [ ] — notation des génotypes : entre parenthèses ( ) Pour une cellule diploïde, les deux allèles sont séparés par une double barre, chaque barre représentant un chromosome homologue Cas de gènes indépendants Cas de gènes liés (bien représenter du même côté les allèles qui sont associés sur un même chromosome) — notation des garnitures chromosomiques Cellule haploïde : n = …Cellule diploïde : 2n = …… correspond au nombre total de chromosomes dans la cellule (qu’ils soient simples ou doubles !)Exemple : – cellule somatique humaine : 2n = 46– gamète humain : n = 23Exercices — vocabulaire de génétique - lignée pure = population dont tous les individus sont homozygotes pour le gène étudié (=> le phénotype ne varie pas de génération en génération) - génération F1 = hybrides F1 = population issue du croisement de deux lignées pures (=> 100 % hétérozygotes) |
||
|
À faire
pour le
15-09-2023
|
Revoir la méiose (et l’alphabet grec)
|
||
|
Commentaires
|
|||
|
Date
|
Vendredi 08 septembre 2023 |
Matière
|
SVT_THOUAND 19
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
Évaluation Test sommaire sur l’alphabet grec Cours ——II.1 Les lois de Mendel [suite]
Exemple : p. 37 doc. 4 Croisement de pois à grains lisses avec des pois à grains ridés -> génération F1 où 100 % [lisses] Croisement de pois F1 avec des pois F1 -> génération F2 où 75 % [lisses] et 25 % [ridés] Ces observations réfutent la théorie de l’hérédité par mélange (on n’obtient pas de pois à grains un peu ridés). Ces observations peuvent être expliquées en supposant : — que chaque parent possède deux facteurs héréditaires — que chaque gamète ne reçoit qu’un seul de ces facteurs héréditaires, aléatoirement — qu’un caractère peut être récessif, un autre dominant (celui qui apparaît seul à la génération F1) -> « Lois de Mendel » (cf. p. 37 doc. 5) Les travaux de Mendel seront oubliés, puis redécouverts quand au XXe siècle, après la découverte des chromosomes, de la méiose et de la fécondation, puis du rôle et de la structure de l’ADN. On comprend alors que les « facteurs héréditaires » de Mendel sont les gènes, avec différents allèles, portés par les chromosomes Annexe au chapitre 1.1 : notions fondamentales de génétique
— un gène -> une protéine -> un caractère (cf. transcription, traduction = étapes de l’expression d’un gène)— un chromosome = une molécule d’ADN (condensée) un gène = segment d’une molécule d’ADN (= segment d’un chromosome) locus d’un gène = position d’un gène sur un chromosome Après réplication de l’ADN -> chromosome double (2 molécules d’ADN IDENTIQUES = chromatides, attachées au niveau du centromère) — chromosomes homologues = chromosomes de même taille, structure, portant les mêmes gènes, mais pas forcément les mêmes allèles = chromosomes « d’une même paire » Pour un gène donné, si les deux chromosomes homologues portent les mêmes allèles, la cellule est dite homozygote ; s’ils portent des allèles différents, la cellule est dite hétérozygote. Notation des génotypes : (A//A) (A//o) Exemple : gène du groupe sanguin, porté par les chromosomes 9 chez l’humain – un individu de génotype (A//A) est homozygote, de groupe sanguin [A] – un individu de génotype (A//o) est hétérozygote, de groupe sanguin [A] => l’allèle o est récessif tandis que l’allèle A est dominant |
||
|
À voir
|
|
||
|
À faire
pour le
13-09-2023
|
— apprendre tout l’alphabet grec (d’alpha à omega)
— revoir la méiose (cours de première, cf documents ci-dessus) |
||
|
Commentaires
|
|||
|
Date
|
Jeudi 07 septembre 2023 |
Matière
|
SVT_THOUAND 21
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
Cours I.1.b Reproduction asexuée des végétaux [suite]
Cf. article « Dans l’Utah, le plus grand être vivant du monde est en danger » (rubrique À voir ci-dessous) I.1.c Développement des individus pluricellulaires
(+ renouvellement continu des tissus : p. 33 doc. 5)Individu pluricellulaire = clone cellulaire de la cellule-œuf = zygote Exercice : Sachant qu’un humain est formé d’environ [latex]\mathrm{3\times 10^{13}}[/latex] cellules, combien au moins y a-t-il eu de divisions depuis le zygote ? [latex]\mathrm{2^n = 3\times 10^{13}}[/latex] [latex]\mathrm{\Leftrightarrow ln(2^n) = ln(3\times 10^{13})}[/latex] [latex]\mathrm{\Leftrightarrow n\times ln(2) = ln(3\times 10^{13})}[/latex] [latex]\mathrm{\Leftrightarrow n = \frac{ln(3\times 10^{13})}{ln(2)}}[/latex] [latex]\mathrm{\Rightarrow}[/latex] Il a fallu au moins 45 mitoses successives pour former un humain adulte à partir du zygote. I.1.d Sélection et amplification clonales des lymphocytes
p. 33 doc. 4 / cours de premièreNombreuses mitoses (amplification) des lymphocytes spécifiques de l’antigène (sélection) => clone cellulaire : grand nombre de lymphocytes présentant la même spécificité que les quelques lymphocytes sélectionnés I.2 Évolution clonale
La diversité génétique dans un clone ne peut résulter que de mutations.Une mutation est à l’origine d’un sous-clone (= lignée cellulaire dérivant de la cellule mutée). Exemples Bactéries
Pas de reproduction sexuée, mais de la diversité génétique grâce aux mutations (la rareté des mutations est compensée par le très grand nombre de mitoses subies) Individus pluricellulaires
Humain : ~ 1 mutation par division cellulaire (cf. p. 35 doc. 6)=> chaque cellule présente au moins 45 mutations par rapport à la cellule-œuf (cf. article « Les vrais jumeaux n’existent pas » ci-dessous) Cancers
Cf. p. 34 doc. 1,2,3Grâce à la protéine P53, les cellules ayant accumulé trop de mutations ne se divisent pas. Mais si c’est le gène p53 qui est muté dans une cellule, ce processus ne fonctionne pas => clone cellulaire dont les cellules accumulent les mutations sans contrôle ; si l’une de ces cellules acquiert une mutation accélérant ses divisions, elle forme une tumeur. —II Le brassage des génomes à chaque génération lors de la reproduction sexuée des individus
——II.1 Les lois de Mendel
Mendel : moine tchèque du XIXe siècle (=> ADN, chromosomes, gènes inconnus, tout comme la méiose et la fécondation)pp. 36-37 : expériences de croisement de pois, afin de comprendre comment sont transmis les caractères héréditaires d’une génération à l’autre Exemple : p. 37 doc. 4 Croisement de pois à grains lisses avec des pois à grains ridés -> génération F1 où 100 % [lisses] Croisement de pois F1 avec des pois F1 -> génération F2 où 75 % [lisses] et 25 % [ridés] Alphabet grec |
||
|
À voir
|
|||
|
À faire
pour le
08-09-2023
|
— apprendre le début de l’alphabet grec (jusqu’à iota). Connaître les lettres minuscules, majuscules et les transcriptions en français
|
||
|
Commentaires
|
|||
|
Date
|
Mercredi 06 septembre 2023 |
Matière
|
SVT_THOUAND 22
|
|
Chapitre
|
1.1 L’origine du génotype des individus
|
Séquence
|
|
|
Résumé
|
Présentation de l’année Enjeux de l’année de terminale : dernière année de lycée
Questionnaire de rentrée
Présentation des épreuves
Épreuve écrite /15 (3 h 30)Épreuve de TP /5 (1 h) Présentation des deux exercices de l’épreuve écrite, et de la grille de notation Moyen de communication
mélhttp://louise.michel.g.free.fr https://agora.louisematic.site (Mattermost du lycée = outil de communication collaborative sur serveur dédié) Cours Thème 1 — Génétique et évolution
Chapitre 1 — L’origine du génotype des individus
Problématique : comment expliquer que la reproduction sexuée forme des individus aux génotypes différents (c’est-à-dire est source de diversité génétique ? I La conservation des génomes : stabilité génétique et évolution clonale
I.1 Clone cellulaire
Succession de mitoses -> production d’un clone = ensemble de cellules toutes génétiquement identiques1 cellule subissant n mitoses : produit [latex]2^n[/latex] cellules Exemples I.1.a Reproduction asexuée des bactéries
Pas de reproduction sexuée chez les procaryotesUne mitose toutes les demi-heures (en conditions optimales) => en 24 h, 1 bactérie peut former [latex]2^{48}[/latex] cellules (= 280 000 milliards) I.1.b Reproduction asexuée des végétaux
cf. bouturage, marcottage |
||
|
À faire
pour le
07-09-2023
|
— s’inscrire sur https://agora.louisematic.site (lien sur la page d’accueil de louisematic). Rejoindre l’équipe Louise Michel, puis le canal de la classe (vous pouvez également rejoindre l’équipe des anciens élèves, si vous souhaitez échanger au sujet de l’orientation)
|
||
|
Commentaires
|
|||